
acizi grași sunt macromolecule organice derivate din hidrocarburi, care sunt compuse din lanțuri lungi de atomi de carbon și hidrogen care sunt hidrofobe (sunt solubili în grăsimi) și reprezintă baza structurală a grăsimilor și lipidelor.
Sunt molecule foarte diverse, care se disting între ele prin lungimea lanțurilor lor de hidrocarburi și prezența, numărul, poziția și / sau configurația legăturilor lor duble..
În lipidele animalelor, plantelor, ciupercilor și microorganismelor, cum ar fi bacteriile și drojdiile, au fost descrise mai mult de 100 de clase diferite de acizi grași și sunt considerate specii și țesuturi specifice la majoritatea ființelor vii..
Uleiurile și grăsimile pe care omul le consumă zilnic sunt de origine animală sau vegetală, sunt compuse în principal din acizi grași.
Indice articol

Moleculele de acizi grași îndeplinesc funcții importante la nivel celular, ceea ce le face componente esențiale și, deoarece unele dintre ele nu pot fi sintetizate de animale, trebuie să le obțină din dietă.
Acizii grași sunt neobișnuiți ca specii libere în citosolul celular, deci se găsesc în general ca parte a altor conjugați moleculari, cum ar fi:
- Lipidele, în membranele biologice.
- Trigliceridele sau esterii acizilor grași, care servesc drept rezerve la plante și animale.
- Ceară, care este ester solid al acizilor grași cu lanț lung și al alcoolilor.
- Alte substanțe similare.
La animale, acizii grași sunt depozitați în citoplasma celulelor sub formă de picături mici de grăsime compuse dintr-un complex numit triacilglicerol, care nu este altceva decât o moleculă de glicerol de care a legat, în fiecare dintre atomii săi de carbon, un lanț de acizi grași prin legături esterice.
În timp ce bacteriile posedă acizi grași scurți și de obicei monoinsaturați, în natură este comun să se găsească acizi grași ale căror lanțuri au un număr par de atomi de carbon, de obicei între 14 și 24, saturați, mononesaturați sau polinesaturați..
Acizii grași sunt molecule amfipatice, adică au două regiuni definite chimic: o regiune polară hidrofilă și o regiune apolară hidrofobă..
Regiunea hidrofobă este alcătuită dintr-un lanț lung de hidrocarburi care, din punct de vedere chimic, nu este foarte reactiv. Regiunea hidrofilă, pe de altă parte, este compusă dintr-o grupare carboxil terminală (-COOH), care se comportă ca un acid.
Această grupă carboxil terminală sau acidul carboxilic ionizează în soluție, este foarte reactivă (din punct de vedere chimic vorbind) și este foarte hidrofilă, reprezentând astfel un loc de legare covalentă între acidul gras și alte molecule..
Lungimea lanțurilor de hidrocarburi ale acizilor grași are de obicei un număr par de atomi de carbon și acest lucru este strâns legat de procesul biosintetic prin care sunt produși, deoarece creșterea lor are loc în perechi de carboni..
Cei mai comuni acizi grași au lanțuri cuprinse între 16 și 18 atomi de carbon, iar la animale, aceste lanțuri nu sunt ramificate.
Acizii grași sunt clasificați în două grupe mari în funcție de natura legăturilor care le compun, adică în funcție de prezența unor legături simple sau duble între atomii de carbon ai lanțurilor lor de hidrocarburi..
Astfel, există acizi grași saturați și nesaturați.
- Acizii grași saturați au legături unice carbon-carbon și toți atomii lor de carbon sunt „saturați” sau atașați la moleculele de hidrogen..
- Acizii grași nesaturați au una sau mai multe duble legături carbon-carbon și nu toți aceștia sunt atașați la un atom de hidrogen.
Acizii grași nesaturați sunt, de asemenea, împărțiți în funcție de numărul de nesaturații (duble legături) în mononesaturați, cei cu o singură legătură dublă și polinesaturați, cei cu mai mult de o.
Au de obicei între 4 și 26 de atomi de carbon legați prin legături simple. Punctul său de topire este direct proporțional cu lungimea lanțului, adică cu greutatea sa moleculară.
Acizii grași care au între 4 și 8 atomi de carbon sunt lichizi la 25 ° C și sunt cei care alcătuiesc uleiuri comestibile, în timp ce cei cu mai mult de 10 atomi de carbon sunt solizi.
Printre cele mai frecvente se numără acidul lauric, care este abundent în uleiurile de miez de palmier și nucă de cocos; acid palmitic, care se găsește în palmier, cacao și untură, și acid stearic, care se găsește în cacao și în uleiurile hidrogenate.
Sunt acizi grași cu mult mai multă stabilitate decât acizii grași nesaturați, în special împotriva oxidării, cel puțin în condiții fiziologice.
Deoarece legăturile unice carbon-carbon se pot roti liber, acizii grași saturați sunt molecule foarte flexibile, deși obstacolele sterice fac ca structura complet extinsă să fie cea mai stabilă din punct de vedere energetic..
Acești acizi grași sunt foarte reactivi și predispuși la saturație și oxidare. Sunt frecvente la plante și la organismele marine. Cei cu o singură legătură dublă sunt cunoscuți ca mononesaturați sau monoenoici, în timp ce cei cu mai mult de doi sunt cunoscuți ca polienoici sau polinesaturați..
Prezența legăturilor duble este comună între atomii de carbon dintre pozițiile 9 și 10, dar acest lucru nu înseamnă că nu se găsesc acizi grași mononesaturați cu o nesaturare în altă poziție.
Spre deosebire de cele saturate, acizii grași nesaturați nu sunt incluși în grupul carboxil terminal, ci în funcție de poziția primei duble legături C - C. Astfel, sunt împărțiți în două grupe, acizii omega-6 sau ω6 și omega-3. sau ω3.
Acizii omega-6 au prima legătură dublă la numărul de carbon 6 și acizii omega-3 au la numărul de carbon 3. Numele ω este dat de legătura dublă cea mai apropiată de grupa metil finală.
Legăturile duble pot fi găsite și în două configurații geometrice cunoscute sub numele de „cis " Da "trans".
Majoritatea acizilor grași nesaturați naturali au configurație „Cis”Iar legăturile duble ale acizilor grași prezenți în grăsimile comerciale (hidrogenate) se găsesc în "trans".
În acizii grași polinesaturați, două legături duble sunt de obicei separate una de cealaltă de cel puțin o grupare metil, adică un atom de carbon legat de doi atomi de hidrogen..
Acizii grași au funcții multiple în organismele vii și, așa cum am menționat mai sus, una dintre funcțiile lor esențiale este ca o parte esențială a lipidelor, care sunt componentele principale ale membranelor biologice și una dintre cele trei cele mai abundente biomolecule din organisme. proteine și carbohidrați.
Sunt, de asemenea, substraturi energetice excelente datorită cărora se obțin cantități mari de energie sub formă de ATP și alți metaboliți intermediari..
Având în vedere că animalele, de exemplu, nu sunt capabile să stocheze carbohidrați, acizii grași reprezintă principala sursă de stocare a energiei care provine din oxidarea zaharurilor consumate în exces.
Acizii grași saturați cu lanț scurt din colon participă la stimularea absorbției apei și a ionilor de sodiu, clorură și bicarbonat; În plus, au funcții în producerea de mucus, în proliferarea colonocitelor (celulelor colonului) etc..
Acizii grași nesaturați sunt abundenți în special în uleiurile vegetale comestibile, care sunt importante în dieta tuturor ființelor umane.

Alții participă ca liganzi ai unor proteine cu activități enzimatice, deci sunt importanți în ceea ce privește efectele lor asupra metabolismului energetic al celulelor în care se găsesc..
Degradarea acizilor grași este cunoscută sub numele de β-oxidare și apare în mitocondriile celulelor eucariote. Dimpotrivă, biosinteza apare în citosolul celulelor animale și în cloroplastele (organite fotosintetice) ale celulelor vegetale..
Este un proces dependent de acetil-CoA, malonil-CoA și NADPH, apare la toate organismele vii și la animale „superioare”, cum ar fi mamiferele. De exemplu, este foarte important în ficatul și țesuturile adipoase, precum și în glandele mamare..
NADPH utilizat pentru această cale este în principal produsul reacțiilor de oxidare dependente de NADP ale căii pentozfosfatului, în timp ce acetil-CoA poate proveni din diferite surse, de exemplu, din decarboxilarea oxidativă a piruvatului, din ciclul Krebs și β-oxidare de acizi grași.
Calea biosintezei, ca și cea a β-oxidării, este extrem de reglementată în toate celulele de către efectori alosterici și modificări covalente ale enzimelor care participă la reglare..
Calea începe cu formarea unui intermediar metabolic cunoscut sub numele de malonil-CoA dintr-o moleculă de acetil-CoA și este catalizat de o enzimă multifuncțională numită acetil-CoA carboxilază..
Această reacție este o reacție de adăugare a unei molecule carboxil dependente de biotină (-COOH, carboxilare) și are loc în două etape:
La animale, formarea lanțurilor de carbohidrați a acizilor grași are loc în continuare prin reacții secvențiale de condensare catalizate de o enzimă multimerică și multifuncțională cunoscută sub numele de acizi grași sintaza..
Această enzimă catalizează condensarea unei unități de acetil-CoA și a mai multor molecule de malonil-CoA care sunt produse din reacția acetil-CoA carboxilază, proces în timpul căruia se eliberează o moleculă de CO2 pentru fiecare malonil-CoA pe care o adaugă.
Acizii grași în creștere sunt esterificați la o proteină numită "proteină purtătoare de acil" sau ACP, care formează tioesterii cu grupări acil. Pe E coli această proteină este o polipeptidă de 10 kDa, dar la animale face parte din complexul de acizi grași sintaza.
Spargerea acestor legături tioester eliberează cantități mari de energie, ceea ce face posibilă, termodinamic vorbind, apariția etapelor de condensare pe calea biosintetică..
La bacterii, activitatea acidului gras sintază corespunde de fapt șase enzime independente care utilizează acetil-coA și malonil-coA pentru a forma acizi grași și cu care sunt asociate șase activități enzimatice diferite.
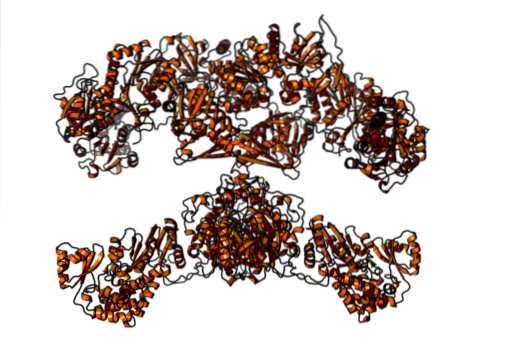
La mamifere, în schimb, acidul gras sintază este un complex enzimatic multifuncțional homodimeric cu o greutate moleculară de aproximativ 500 kDa, care are șase activități catalitice diferite și cu care proteina purtătoare de acil se asocia.
Grupurile tiol din reziduurile de cisteină responsabile de legarea intermediarilor metabolici de enzima ACP trebuie încărcate, înainte de începerea sintezei, cu grupările acil necesare..
Pentru aceasta, grupul acetil al acetil-coA este transferat la gruparea tiol (-SH) a unuia dintre resturile de cisteină ale subunității ACP a sintazei acizilor grași. Această reacție este catalizată de subunitatea ACP-acil-transferază..
Gruparea acetil este apoi transferată de la ACP la un alt reziduu de cisteină la locul catalitic al unei alte subunități enzimatice a complexului cunoscut sub numele de β-cetoacil-ACP-sintază. Astfel, complexul enzimatic este „pregătit” pentru a începe sinteza..
Malonil-CoA care este produs de acetil-CoA carboxilază este transferat la gruparea tiol în ACP și în timpul acestei reacții se pierde porțiunea CoA. Reacția este catalizată de subunitatea malonil-ACP-transferază a complexului de acid gras sintază, care produce apoi malonil-ACP.
În timpul acestui proces, gruparea malonil este legată de ACP și β-cetoacil-ACP-sintază printr-un ester și, respectiv, o altă legătură sulfhidril..
Enzima β-cetoacil-ACP-sintază catalizează transferul grupării acetil care i-a fost atașată în etapa de „amorsare” la 2-carbonul grupării malonil care, în etapa anterioară, a fost transferat la ACP.
În timpul acestei reacții, o moleculă de CO2 este eliberată din malonil, care corespunde CO2 emis de bicarbonat în reacția de carboxilare a acetil-CoA carboxilazei. Acetoacetil-ACP este apoi produs.
Subunitatea β-cetoacil-ACP-reductază catalizează reducerea dependentă de NADPH a acetoacetil-ACP, formând astfel D-β-hidroxibutiril-ACP.
În această etapă, se formează trans-α, β-acil-ACP sau ∆2-nesaturat-acil-ACP (cratonil-ACP), produs al deshidratării D-β-hidroxibutiril-ACP prin acțiunea enoil- subunitate.ACP-hidratază.
Mai târziu, cratonil-ACP este redus la butiril-ACP printr-o reacție dependentă de NADPH catalizată de subunitatea enil-ACP-reductază. Această reacție finalizează primul din cele șapte cicluri necesare pentru a produce palmitoil-ACP, care este un precursor al aproape tuturor acizilor grași..
Gruparea butiril este transferată de la ACP la gruparea tiol a unui reziduu de cisteină în β-cetoacil-ACP-sintază, cu care ACP este capabil să accepte o altă grupare malonil din malonil-CoA.
În acest fel, reacția care apare este condensarea malonil-ACP cu buturil-β-cetoacil-ACP-sintază, ceea ce dă naștere la β-cetohexanoil-ACP + CO2.
Palmitoil-ACP care apare din etapele ulterioare (după adăugarea a încă 5 unități malonil) poate fi eliberat ca acid palmitic liber datorită activității enzimei tioesterazice, poate fi transferat la CoA sau încorporat în acid fosfatidic pentru fosfolipid și calea de sinteză a triacilgliceridelor.
Sinteza acizilor grași a majorității organismelor se oprește în sinteza palmitoil-ACP, deoarece situl catalitic al subunității β-cetoacil-ACP-sintază are o configurație în care numai acizii grași de acea lungime pot fi găzduiți.
Acestea sunt relativ frecvente în organismele marine și sunt sintetizate și de un complex de acizi grași sintaza. Cu toate acestea, reacția de „amorsare” are loc cu o moleculă mai lungă, propionil-ACP, cu trei atomi de carbon..
Acidul palmitic, după cum sa discutat, servește ca un precursor pentru mulți acizi grași saturați și nesaturați cu lanț mai lung. Procesul de „alungire” a acizilor grași are loc în mitocondrii, în timp ce introducerea nesaturărilor are loc în esență în reticulul endoplasmatic.
Multe organisme își convertesc acizii grași saturați în acizi nesaturați ca o adaptare la temperaturi scăzute ale mediului, deoarece acest lucru le permite să mențină punctul de topire a lipidelor sub temperatura camerei..
Multe dintre proprietățile acizilor grași depind de lungimea lanțului lor și de prezența și numărul de nesaturații:
- Acizii grași nesaturați au puncte de topire mai mici decât acizii grași saturați de aceeași lungime.
- Lungimea acizilor grași (numărul de atomi de carbon) este invers proporțională cu fluiditatea sau flexibilitatea moleculei, adică moleculele „mai scurte” sunt mai fluide și invers..
În general, substanțele grase fluide sunt compuse din acizi grași cu lanț scurt cu prezența nesaturărilor.
Plantele au cantități abundente de acizi grași nesaturați, precum și animale care trăiesc la temperaturi foarte scăzute, deoarece acestea, ca componente ale lipidelor prezente în membranele celulare, le conferă o fluiditate mai mare în aceste condiții..
În condiții fiziologice, prezența unei duble legături în lanțul hidrocarbonat al unui acid gras determină o curbură de aproximativ 30 °, ceea ce face ca aceste molecule să ocupe un spațiu mai mare și să scadă rezistența interacțiunilor lor van der Waals..
Prezența legăturilor duble în acizii grași asociați cu moleculele lipidice are efecte directe asupra gradului de „ambalare” pe care acestea îl pot avea în membranele la care aparțin și, astfel, au efecte și asupra proteinelor de membrană.
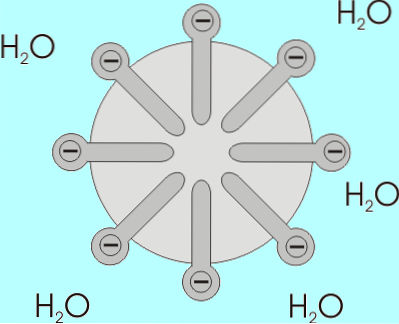
Solubilitatea acizilor grași scade odată cu creșterea lungimii lanțului lor, deci sunt invers proporționale. În amestecurile apoase și lipidice, acizii grași se asociază în structuri cunoscute sub numele de micele..
O micelă este o structură în care lanțurile alifatice ale acizilor grași sunt „închise”, astfel „expulzând” toate moleculele de apă și pe a căror suprafață se găsesc grupările carboxil..
Nomenclatura acizilor grași poate fi oarecum complexă, mai ales dacă se referă la denumirile comune pe care le primesc, care sunt adesea legate de unele proprietăți fizico-chimice, cu locul în care se găsesc sau alte caracteristici.
Mulți autori consideră că, deoarece aceste molecule sunt ionizate la pH fiziologic datorită grupării carboxil terminale, ar trebui să le numim „carboxilați” folosind terminația „ato ".
Conform sistemului IUPAC, enumerarea atomilor de carbon ai unui acid gras se face din grupa carboxil la capătul polar al moleculei și primii doi atomi de carbon atașați la această grupă sunt numiți α și respectiv β. Metilul terminal al lanțului conține atomul de carbon ω.
În general, în nomenclatura sistematică li se dă numele hidrocarburii „părinte” (hidrocarbura cu același număr de atomi de carbon) și terminarea acesteia „sau" pentru „Oico”, dacă este un acid gras nesaturat, se adaugă finalul „Enoic”.
Luați în considerare, de exemplu, cazul unui acid gras C18 (C18):
- Deoarece hidrocarbura cu același număr de atomi de carbon este cunoscută sub numele de octadecan, acidul saturat se numește „acid octadecanoic"O bine"octadecanoat”Și numele său comun este acid stearic.
- Dacă aveți o legătură dublă între unele perechi de atomi de carbon din structura dvs., este cunoscut sub numele de „acid octadecenoic"
- Dacă are două legături duble c - c, atunci se numește „Acid octadecadienoic” si daca ai trei "acid octadecatrienoic".
Dacă doriți să rezumați nomenclatura, atunci 18: 0 este utilizat pentru acidul gras 18-carbon și fără duble legături (saturate) și, în funcție de gradul de nesaturare, atunci în loc de zero, 18: 1 este scris pentru o moleculă cu o nesaturare, 18: 2 pentru una cu două nesaturări și așa mai departe.
Dacă doriți să specificați între ce atomi de carbon sunt legăturile duble din acizii grași nesaturați, utilizați simbolul ∆ cu un supernumeric numeric care indică locul nesaturării și prefixul „Cis"Sau "trans", în funcție de configurația acestuia.


Nimeni nu a comentat acest articol încă.