enzime de restricție sunt endonucleaze utilizate de anumite arhee și bacterii pentru a inhiba sau „restricționa” răspândirea virusurilor în interiorul lor. Sunt deosebit de frecvente la bacterii și fac parte din sistemul lor de apărare împotriva ADN-ului străin cunoscut sub numele de sistem de restricție / modificare..
Aceste enzime catalizează scindarea ADN-ului cu bandă dublă în locații specifice, reproductibil și fără utilizarea energiei suplimentare. Majoritatea necesită prezența cofactorilor precum magneziu sau alți cationi divalenți, deși unii necesită și ATP sau S-adenosil metionină.
Endonucleazele de restricție au fost descoperite în 1978 de Daniel Nathans, Arber Werner și Hamilton Smith, care au primit premiul Nobel pentru medicină pentru descoperirea lor. Numele lor derivă în general din organismul în care sunt observați pentru prima dată.
Astfel de enzime sunt utilizate pe scară largă în dezvoltarea metodelor de clonare a ADN-ului și a altor strategii de biologie moleculară și inginerie genetică. Caracteristicile lor specifice de recunoaștere a secvenței și capacitatea de a tăia secvențe aproape de siturile de recunoaștere le fac instrumente puternice în experimentarea genetică..
Fragmentele generate de enzimele de restricție care au acționat asupra unei anumite molecule de ADN pot fi folosite pentru a recrea o „hartă” a moleculei originale utilizând informații despre siturile în care enzima a tăiat ADN-ul..
Unele enzime de restricție pot avea același loc de recunoaștere pe ADN, dar nu îl taie neapărat în același mod. Astfel, există enzime care taie capetele contondente lăsate și enzime care taie capetele coezive, care au aplicații diferite în biologia moleculară.
În prezent, există sute de enzime de restricție disponibile în comerț, oferite de diferite case comerciale; aceste enzime funcționează ca foarfece moleculare „personalizate” în scopuri diferite.
Indice articol
Enzimele de restricție îndeplinesc funcția opusă a polimerazelor, deoarece hidrolizează sau rup legătura esterică în legătura fosfodiesterică dintre nucleotidele adiacente într-un lanț nucleotidic..
În biologia moleculară și ingineria genetică, acestea sunt instrumente utilizate pe scară largă pentru construirea vectorilor de expresie și clonare, precum și pentru identificarea secvențelor specifice. Ele sunt, de asemenea, utile pentru construirea genomilor recombinați și au un mare potențial biotehnologic..
Progresele recente în terapia genică utilizează în prezent enzime de restricție pentru introducerea anumitor gene în vectori care sunt vehicule pentru transportul unor astfel de gene în celule vii și care probabil au capacitatea de a se introduce în genomul celular pentru a efectua modificări permanente.
Enzimele de restricție pot cataliza scindarea ADN-ului cu bandă dublă, deși unele sunt capabile să recunoască secvențe ADN cu bandă unică și chiar ARN. Tăierea are loc după recunoașterea secvențelor.
Mecanismul de acțiune constă în hidroliza legăturii fosfodiesterice dintre o grupare fosfat și o dezoxiriboză din coloana vertebrală a fiecărei catene de ADN. Multe dintre enzime sunt capabile să taie în același loc pe care îl recunosc, în timp ce altele taie între 5 și 9 perechi de baze înainte sau după aceeași.
În mod normal, aceste enzime sunt tăiate la capătul 5 'al grupării fosfat, dând naștere la fragmente de ADN cu capătul 5' fosforil și capătul hidroxil terminal 3 '..
Deoarece proteinele nu intră în contact direct cu situl de recunoaștere din ADN, acestea trebuie translocate succesiv până când se realizează situsul specific, probabil prin intermediul mecanismelor de „alunecare” pe catena de ADN..
În timpul clivajului enzimatic, legătura fosfodiesterică a fiecăreia dintre firele de ADN este poziționată într-unul dintre locurile active ale enzimelor de restricție. Când enzima părăsește situl de recunoaștere și decolteu, o face prin asocieri tranzitorii nespecifice.
În prezent sunt cunoscute cinci tipuri de enzime de restricție. Iată o scurtă descriere a fiecăruia:
Aceste enzime sunt proteine pentamerice mari, cu trei subunități, una pentru restricție, una pentru metilare și una pentru recunoașterea secvenței în ADN. Aceste endonucleaze sunt proteine multifuncționale capabile să catalizeze reacții de restricție și modificare, au activitate ATPază și, de asemenea, topoizomerază ADN..
Enzimele de acest tip au fost primele endonucleaze descoperite, au fost purificate pentru prima dată în anii 1960 și au fost studiate în profunzime încă de atunci.
Enzimele de tip I nu sunt utilizate pe scară largă ca instrument biotehnologic, deoarece situl de scindare poate fi la o distanță variabilă de până la 1.000 de perechi de baze de la locul de recunoaștere, ceea ce le face nesigure în ceea ce privește reproductibilitatea experimentală..
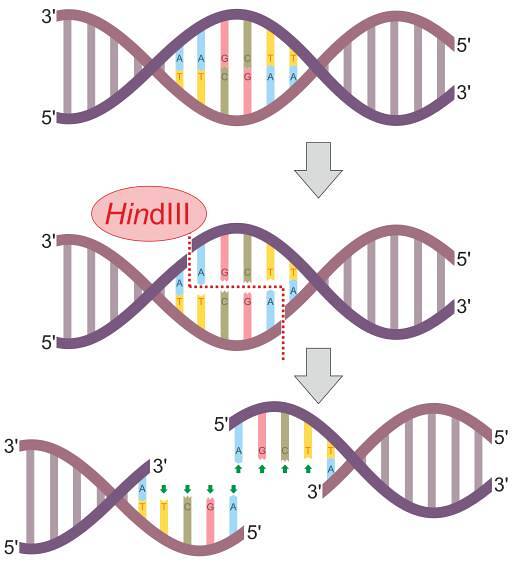
Sunt enzime compuse din homodimeri sau tetrameri care taie ADN-ul în locuri definite între 4 și 8 bp în lungime. Aceste site-uri de scindare sunt de obicei palindromice, adică recunosc secvențe care sunt citite în același mod în ambele direcții..
Multe dintre enzimele de restricție de tip II din bacterii taie ADN-ul atunci când recunosc caracterul său străin, deoarece nu are modificările tipice pe care propriul ADN ar trebui să le aibă.
Acestea sunt cele mai simple enzime de restricție, deoarece nu necesită alt cofactor decât magneziul (Mg +) pentru a recunoaște și tăia secvențele ADN..
Precizia enzimelor de restricție de tip II în recunoașterea și tăierea secvențelor simple din ADN în poziții precise le face una dintre cele mai utilizate și indispensabile în majoritatea ramurilor biologiei moleculare..
În cadrul grupului de enzime de restricție de tip II există mai multe subclase clasificate în funcție de anumite proprietăți care sunt unice pentru fiecare. Clasificarea acestor enzime se face prin adăugarea literelor alfabetului, de la A la Z urmând denumirea enzimei.
Unele dintre subclasele cele mai cunoscute pentru utilitatea lor sunt:
Sunt dimeri ai diferitelor subunități. Ele recunosc secvențe asimetrice și sunt utilizate ca precursori ideali pentru generarea enzimelor de tăiere.
Acestea sunt alcătuite din unul sau mai mulți dimeri și taie ADN pe ambele părți ale secvenței de recunoaștere. Tăiați ambele fire de ADN la un interval de perechi de baze în fața locului de recunoaștere.
Enzimele de acest tip sunt polipeptide cu funcții de diviziune și modificare a catenelor de ADN. Aceste enzime taie ambele fire asimetric.
Enzimele acestei subclase sunt cele mai utilizate în ingineria genetică. Au un situs catalitic și necesită în general un efector alosteric. Aceste enzime trebuie să interacționeze cu două copii ale secvenței lor de recunoaștere pentru a efectua un clivaj eficient. În această subclasă se află enzimele EcoRII și EcoRI.
Endonucleazele de restricție de tip III sunt compuse din doar două subunități, una este responsabilă pentru recunoașterea și modificarea ADN, în timp ce cealaltă este responsabilă pentru scindarea secvenței.
Aceste enzime necesită doi cofactori pentru funcția lor: ATP și magneziu. Enzimele de restricție de acest tip posedă două situri de recunoaștere asimetrice, translocează ADN într-o manieră dependentă de ATP și îl taie între 20 și 30 pb adiacent situsului de recunoaștere..
Enzimele de tip IV sunt ușor de identificat, deoarece taie ADN-ul cu semne de metilare, sunt alcătuite din mai multe subunități diferite care sunt responsabile pentru recunoașterea și tăierea secvenței ADN. Aceste enzime folosesc GTP și magneziu divalent ca cofactori.
Siturile de clivaj specifice includ fire de nucleotide cu reziduuri de citozină metilate sau hidroximetilate pe una sau ambele fire de acizi nucleici..
Această clasificare grupează împreună enzimele de tip CRISPER-Cas, care identifică și taie secvențe specifice de ADN de la organismele invadatoare. Enzimele Cas utilizează un fir de ARN ghid sintetizat de la CRISPER pentru a recunoaște și a ataca organismele invadatoare.
Enzimele clasificate ca tip V sunt polipeptide structurate prin enzime de tip I, II și II. Pot tăia secțiuni din ADN-ul a aproape orice organism și cu o gamă largă de lungime. Flexibilitatea și ușurința lor de utilizare fac din aceste enzime unul dintre cele mai utilizate instrumente în ingineria genetică în prezent, alături de enzimele de tip II..
Enzimele de restricție au fost utilizate pentru detectarea polimorfismelor ADN, în special în studii genetice ale populației și studii evolutive care utilizează ADN mitocondrial, pentru a obține informații despre ratele de substituție a nucleotidelor..
În prezent, vectorii utilizați pentru transformarea bacteriilor în diferite scopuri au site-uri de multiclonare în care există site-uri de recunoaștere pentru enzime de restricție multiple..
Printre aceste enzime cele mai populare sunt EcoRI, II, III, IV și V, obținute și descrise pentru prima dată din E coli; HindIII, din H. influenzae și BamHI din B. amyloliquefaciens.


Nimeni nu a comentat acest articol încă.