
glicoliză sau glicoliza este principala cale a catabolismului glucozei, al cărui obiectiv final este de a obține energie sub formă de ATP și reducerea puterii sub formă de NADH, din acest carbohidrat.
Această cale, complet elucidată în anii 1930 de Gustav Embden și Otto Meyerhof în timp ce studia consumul de glucoză în celulele musculare scheletice, constă în oxidarea completă a acestei monozaharide și, prin ea însăși, reprezintă o cale anaerobă pentru glucoză..
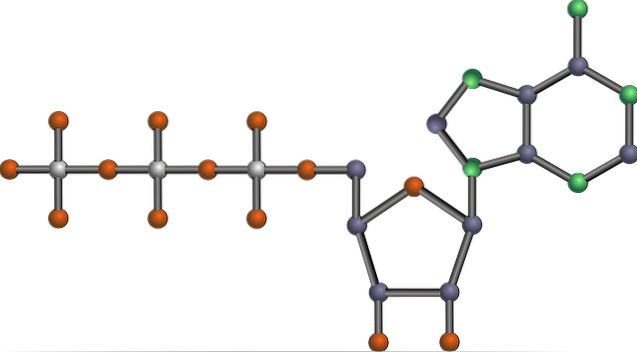
Este una dintre principalele căi metabolice, deoarece apare, cu diferențele sale, în toate organismele vii care există, unicelulare sau multicelulare, procariote sau eucariote și se crede că este un lanț de reacții care este conservat în mod evolutiv în natură..
De fapt, există unele organisme și tipuri de celule care depind exclusiv de această cale pentru a supraviețui..
În primul caz, glicoliza constă în oxidarea glucozei, a 6 atomi de carbon, până la piruvat, care are trei atomi de carbon; cu producerea concomitentă de ATP și NADH, utilă pentru celule din punct de vedere metabolic și sintetic.
În celulele capabile să proceseze în continuare produsele obținute din catabolismul glucozei, glicoliza se încheie cu producerea de dioxid de carbon și apă prin ciclul Krebs și prin lanțul de transport al electronilor (glicoliza aerobă)..
Zece reacții enzimatice au loc în cursul căii glicolitice și, deși reglarea acestor reacții poate fi oarecum diferită de la o specie la alta, mecanismele de reglare sunt, de asemenea, destul de conservate..
Indice articol
Din punct de vedere metabolic, glucoza este unul dintre cei mai importanți carbohidrați pentru toate ființele vii.
Este o moleculă stabilă și foarte solubilă, deci poate fi transportată cu relativă ușurință pe tot corpul unui animal sau al unei plante, de unde este depozitată și / sau obținută până unde este necesară ca combustibil celular..
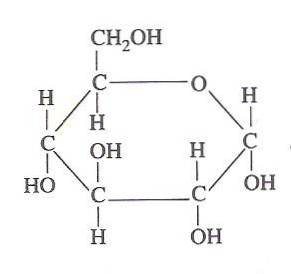
Energia chimică conținută în glucoză este exploatată de celulele vii prin glicoliză, care constă dintr-o serie de etape foarte controlate prin care energia eliberată de oxidarea acestui carbohidrat poate fi „capturată” în forme de energie mai utilizabile., De unde și importanța sa.
Prin această cale, se obțin nu numai energia (ATP) și puterea de reducere (NADH), dar oferă și o serie de intermediari metabolici care fac parte din alte căi, importante și din funcția anabolică (biosintetică) și funcționarea celulară generală. Iată o listă:
- Glucoză 6-fosfat pentru calea pentozei fosfat (PPP) Calea fosfatului pentozei)
- Piruvat pentru fermentarea lactică
- Piruvat pentru sinteza aminoacizilor (alanină, în principal)
- Piruvat pentru ciclul acidului tricarboxilic
- Fructoza 6-fosfat, glucoza 6-fosfat și dihidroxiacetonă fosfat, care funcționează ca „blocuri de construcție” în alte căi precum sinteza glicogenului, acizilor grași, trigliceridelor, nucleotidelor, aminoacizilor etc..
Cantitatea de ATP produsă de calea glicolitică, atunci când celula care o produce nu poate trăi în condiții aerobe, este suficientă pentru a furniza nevoile energetice ale unei celule atunci când este cuplată la diferite tipuri de procese de fermentare.
Cu toate acestea, când vine vorba de celule aerobe, glicoliza servește și ca sursă de energie de urgență și servește ca „etapă pregătitoare” înainte de reacțiile de fosforilare oxidativă care caracterizează celulele cu metabolism aerob..
Glicoliza este posibilă numai datorită participării celor 10 enzime care catalizează reacțiile care caracterizează această cale. Multe dintre aceste enzime sunt alosterice și își schimbă forma sau conformația atunci când își exercită funcțiile catalitice..
Există enzime care se rup și formează legături covalente între substraturile lor și există altele care necesită cofactori specifici pentru a-și îndeplini funcțiile, în principal ioni metalici..
Structural vorbind, toate enzimele glicolitice au un centru esențial laminat. β paralelă inconjurat de helici α și comandate în mai multe domenii. Mai mult, aceste enzime sunt caracterizate prin faptul că siturile lor active sunt de obicei la locurile de legare dintre domenii..
De asemenea, este important să rețineți că principala reglare a traseului trece prin controlul (hormonal sau metaboliți) al enzimelor precum hexokinaza, fosfofructokinaza, gliceraldehida 3-fosfat dehidrogenaza și piruvatul kinaza..

Prima reacție de glicoliză (fosforilarea glucozei) este catalizată de hexokinază (HK), al cărei mecanism de acțiune pare a consta dintr-un substrat „strângere indusă” care promovează „blocarea” enzimei în jurul ATP și a glucozei (substraturile sale) odată s-a legat de acestea.
În funcție de organismul considerat, pot exista una sau mai multe izoenzime, a căror greutate moleculară variază între 50 (aproximativ 500 aminoacizi) și 100 kDa, deoarece acestea par să se grupeze împreună sub formă de dimeri, a căror formare este favorizată de prezența glucozei, ionilor de magneziu și ATP.
Hexokinaza are o structură terțiară compusă din foi alfa și beta deschise, deși există multe diferențe structurale în aceste enzime.
Glucoza fosforilată de hexokinază este izomerizată în fructoză 6-fosfat prin fosfoglucoză izomerază (IGP), cunoscută și sub denumirea de glucoză 6-fosfat izomerază. Enzima, atunci, nu îndepărtează sau adaugă atomi, ci îi rearanjează la nivel structural.
Aceasta este o enzimă activă în forma sa dimerică (monomerul cântărește mai mult sau mai puțin 66 kDa) și este implicat nu numai în glicoliză, ci și în gluconeogeneză, în sinteza carbohidraților în plante etc..
Fructoza 6-fosfat este un substrat pentru enzima fosfofructokinază, care este capabilă să refosforileze această moleculă utilizând ATP ca donator de grup fosforil, producând fructoză 1,6-bisfosfat.
Această enzimă există la bacterii și mamifere ca enzimă homotetramerică (compusă din patru subunități identice de 33 kDa fiecare pentru bacterii și 85 kDa fiecare la mamifere), iar în drojdie este un octamer (compus din subunități mai mari, între 112 și 118 kDa).
Este o enzimă alosterică, ceea ce înseamnă că este reglată pozitiv sau negativ de unii dintre produsele sale (ADP) și de alte molecule precum ATP și citrat..
Cunoscută și sub numele de fructoză 1,6-bisfosfat aldolază, aldolaza catalizează defalcarea catalitică a fructozei 1,6-bisfosfat în dihidroxiacetonă fosfat și gliceraldehidă 3-fosfat și reacția inversă, adică unirea ambelor zaharuri pentru formarea fructozei 1 , 6-bifosfat.
Cu alte cuvinte, această enzimă reduce fructoza 1,6-bifosfat chiar în jumătate, eliberând doi compuși C3 fosforilați. Aldolaza este, de asemenea, compusă din 4 subunități identice, fiecare cu propriul său site activ..
A fost determinată existența a două clase (I și II) ale acestei enzime, care se diferențiază prin mecanismul reacției pe care o catalizează și pentru că unele (primele) apar la bacterii și eucariote „inferioare”, iar celelalte (a doua ) se află în bacterii, protiști și metazoane.
Aldolaza eucariotă „superioară” constă dintr-un homotetramer de subunități cu greutate moleculară de 40 kDa, fiecare constând dintr-un butoi compus din 8 foi β / α..
Cele două trioze fosforilate pot fi interconvertite între ele datorită acțiunii izomerazei trioz-fosfat, care permite utilizarea ambelor zaharuri pe tot parcursul glicolizei, asigurând utilizarea deplină a fiecărei molecule de glucoză care intră pe cale..
Această enzimă a fost descrisă ca enzima „perfectă”, deoarece catalizează reacția descrisă de aproximativ trilioane de ori mai rapid decât s-ar produce fără participarea dumneavoastră. Situl său activ se află în centrul unei structuri beta-barilice, caracteristică multor enzime glicolitice..
Este o proteină dimerică, formată din două subunități identice de aproximativ 27 kDa, ambele cu structură globulară..
Gliceraldehida 3-fosfatul produs prin acțiunea aldolazei și triozei fosfat izomerazei servește ca substrat pentru GAPDH, care este o enzimă homotetramerică (34-38 kDa fiecare subunitate) care se leagă în mod cooperant la o moleculă de NAD + în fiecare dintre cele 4 situsuri active , precum și 2 ioni fosfat sau sulfat.
În această etapă a căii, enzima permite fosforilarea unuia dintre substraturile sale folosind fosfat anorganic ca donator de grup fosforil, cu reducerea concomitentă a două molecule NAD + și producerea 1,3-bisfosfogliceratului..
Fosfogliceratul kinazic este responsabil pentru transferul uneia dintre grupările fosfat de 1,3-bisfosfoglicerat la o moleculă ADP prin fosforilare la nivelul substratului. Această enzimă folosește un mecanism similar cu cel utilizat de hexokinază, deoarece se închide la contactul pe substraturile sale, protejându-le de moleculele de apă care interferează..
Această enzimă, ca și altele care utilizează două sau mai multe substraturi, au un loc de legare pentru ADP și un altul pentru fosfatul de zahăr.
Spre deosebire de celelalte enzime descrise, această proteină este un monomer de 44 kDa cu o structură bilobară, compusă din două domenii de aceeași dimensiune conectate printr-o „despicătură” îngustă..
3-fosfogliceratul suferă o schimbare de la gruparea fosfat la carbonul 2, în mijlocul moleculei, ceea ce reprezintă un loc de instabilitate strategică care facilitează transferul ulterior al grupului către o moleculă ATP în ultima reacție a căii..
Această rearanjare este catalizată de enzima fosfoglicerat mutază, o enzimă dimerică pentru oameni și tetramerică pentru drojdie, cu o dimensiune a subunității apropiată de 27 kDa..
Enolaza catalizează deshidratarea 2-fosfogliceratului în fosfoenolpiruvat, un pas necesar pentru generarea de ATP în reacția ulterioară.
Este o enzimă dimerică compusă din două subunități identice de 45 kDa. Depinde de ioni de magneziu pentru stabilitatea sa și pentru schimbarea conformațională necesară pentru a se lega de substratul său. Este una dintre enzimele exprimate cel mai abundent în citosolul multor organisme și îndeplinește funcții în plus față de glicolitice..
A doua fosforilare la nivel de substrat care are loc în glicoliză este catalizată de piruvat kinază, care este responsabilă pentru transferul grupării fosforil de la fosfoenolpiruvat la ADP și pentru producerea piruvatului..
Această enzimă este mai complexă decât oricare dintre celelalte enzime glicolitice și la mamifere este o enzimă homotetramerică (57 kDa / subunitate). Există, la vertebrate, cel puțin 4 izoenzime: L (în ficat), R (în eritrocite), M1 (în mușchi și creier) și M2 (țesut fetal și țesuturi adulte).
Calea glicolitică constă din zece etape secvențiale și începe cu o moleculă de glucoză. În timpul procesului, molecula de glucoză este „activată” sau „preparată” cu adăugarea a doi fosfați, inversând două molecule de ATP..
Mai târziu este „tăiat” în două fragmente și în cele din urmă este modificat chimic de câteva ori, sintetizând patru molecule ATP pe parcurs, astfel încât câștigul net al traseului să corespundă cu două molecule ATP..
Din cele de mai sus, se poate deduce că ruta este împărțită într-o fază de „investiție” energetică, fundamentală pentru oxidarea completă a moleculei de glucoză, și o altă fază de „câștig” energetic, unde energia utilizată inițial este înlocuită și două ATP nete molecule.
1- Primul pas al căii glicolitice constă în fosforilarea glucozei mediată de hexokinază (HK), pentru care enzima folosește o moleculă de ATP pentru fiecare moleculă de glucoză fosforilată. Este o reacție ireversibilă și depinde de prezența ionilor de magneziu (Mg2 +):
Glucoza + ATP → Glucoza 6-fosfat + ADP
2- Glucoza 6-fosfatul astfel produs este izomerizat în fructoză 6-fosfat datorită acțiunii enzimei fosfoglucoză izomerază (IGP). Aceasta este o reacție reversibilă și nu implică cheltuieli suplimentare de energie:
Glucoză 6-fosfat → Fructoză 6-fosfat
3- Ulterior, o altă etapă de inversare a energiei implică fosforilarea fructozei 6-fosfat pentru a forma fructoză 1,6-bisfosfat. Această reacție este catalizată de enzima fosfofructokinază-1 (PFK-1). La fel ca primul pas în cale, molecula donatoare a grupului fosfat este ATP și este, de asemenea, o reacție ireversibilă.
Fructoză 6-fosfat + ATP → Fructoză 1,6-bisfosfat + ADP
4- În această etapă a glicolizei, există descompunerea catalitică a fructozei 1,6-bisfosfat în dihidroxiacetonă fosfat (DHAP), o cetoză și gliceraldehidă 3-fosfat (GAP), o aldoză. Această condensare a aldolului este catalizată de enzima aldolază și este un proces reversibil..
Fructoză 1,6-bisfosfat → Dihidroxiacetonă fosfat + gliceraldehidă 3-fosfat
5- Ultima reacție a fazei de inversiune a energiei constă în interconversia triosfosfatului DHAP și GAP catalizată de enzima triozofosfat izomerazei (TIM), fapt care nu necesită aport suplimentar de energie și este, de asemenea, un proces reversibil.
Dihidroxiacetonă fosfat ↔ Gliceraldehidă 3-fosfat
6- Gliceraldehida 3-fosfatul este utilizat „în aval” în calea glicolitică ca substrat pentru o reacție de oxidare și altul pentru fosforilare, catalizat de aceeași enzimă, gliceraldehidă 3-fosfat dehidrogenază (GAPDH)..
Enzima catalizează oxidarea carbonului C1 al moleculei la acid carboxilic și fosforilarea acesteia în aceeași poziție, producând 1,3-bisfosfoglicerat. În timpul reacției, 2 molecule de NAD + sunt reduse pentru fiecare moleculă de glucoză și se utilizează 2 molecule de fosfat anorganic..
2Gliceraldehidă 3-fosfat + 2NAD + + 2Pi → 2 (1,3-bisfosfoglicerat) + 2NADH + 2H
În organismele aerobe, fiecare NADH produs în acest fel trece prin lanțul de transport al electronilor pentru a servi ca substrat pentru sinteza a 6 molecule ATP prin fosforilare oxidativă..
7- Aceasta este prima etapă de sinteză a ATP în glicoliză și implică acțiunea fosfoglicerat kinazei (PGK) asupra 1,3-bisfosfogliceratului, transferând o grupare fosforil (fosforilare la nivel de substrat) de la această moleculă la o moleculă de ADP, producând 2ATP și 2 molecule de 3-fosfoglicerat (3PG) pentru fiecare moleculă de glucoză.
2 (1,3-bisfosfoglicerat) + 2ADP → 2 (3-fosfoglicerat) + 2ATP
8- 3-fosfogliceratul servește ca substrat pentru enzima fosfoglicerat mutază (PGM), care îl transformă în 2-fosfoglicerat prin deplasarea grupării fosforil din carbon 3 în carbon 2 printr-o reacție în două etape care este reversibilă și dependentă de ioni de magneziu (Mg + 2).
2 (3-fosfoglicerat) → 2 (2-fosfoglicerat)
9- Enzima enzimă enolazică deshidratează 2-fosfogliceratul și produce fosfoenolpiruvat (PEP) printr-o reacție care nu necesită adăugarea de energie suplimentară și al cărei scop este de a produce un compus cu energie ridicată, capabil să doneze gruparea sa fosforil în următoarea reacție.
2 (2-fosfoglicerat) → 2 Fosfoenolpiruvat
10- Fosfoenolpiruvatul este un substrat pentru enzima piruvat kinaza (PYK), care este responsabilă pentru transferul grupării fosforil din această moleculă la o moleculă ADP, catalizând astfel o altă reacție de fosforilare la nivelul substratului.
În reacție, se produc 2ATP și 2 molecule de piruvat pentru fiecare glucoză și este necesară prezența de potasiu și magneziu sub formă ionică..
2Fosfoenolpiruvat + 2ADP → 2Piruvat + 2ATP
Randamentul net al glicolizei, în acest fel, constă în 2ATP și 2NAD + pentru fiecare moleculă de glucoză care intră pe cale..
Dacă este vorba de celule cu metabolism aerob, atunci degradarea totală a unei molecule de glucoză produce între 30 și 32 ATP prin ciclul Krebs și prin lanțul de transport al electronilor.
Reacția generală a glicolizei este următoarea:
Glucoza + 2NAD + + 2ADP + 2Pi → 2Piruvat + 2ATP + 2NADH + 2H+
Prin urmare, dacă este analizat pe scurt, s-ar putea asigura că principalele produse ale căii glicolitice sunt piruvatul, ATP, NADH și H.
Cu toate acestea, soarta metabolică a fiecărui intermediar de reacție depinde, în mare măsură, de nevoile celulare, motiv pentru care toți intermediarii pot fi considerați ca produse de reacție și pot fi enumerați după cum urmează:
- Glucoză 6-fosfat
- Fructoza 6-fosfat
- Fructoză 1,6-bifosfat
- Dihidroxiacetonă fosfat și gliceraldehidă 3-fosfat
- 1,3-bifosfoglicerat
- 3-fosfoglicerat și 2-fosfoglicerat
- Fosfoenolpiruvat și piruvat
În ciuda faptului că glicoliza, de la sine (se poate vorbi de glicoliză anaerobă), produce doar aproximativ 5% din ATP care poate fi extras din catabolismul aerob al glucozei, această cale metabolică este esențială din mai multe motive:
- Acesta servește ca o sursă „rapidă” de energie, în special în situațiile în care un animal trebuie să iasă rapid dintr-o stare de repaus, pentru care procesele de oxidare aerobă nu ar fi suficient de rapide.
- Fibrele musculare scheletice „albe” din corpul uman, de exemplu, sunt fibre cu contracție rapidă și depind de glicoliza anaerobă pentru a funcționa..
- Când, dintr-un anumit motiv, o celulă trebuie să renunțe la o parte din mitocondriile sale (care sunt organitele care efectuează fosforilarea oxidativă a unei părți a produselor glicolitice, printre altele), celula devine mai dependentă de energia obținută prin glicolit..
- Multe celule depind de glucoză ca sursă de energie prin calea glicolitică, printre care globule rosii, lipsit de organite interne și celule ale ochiului (în special cele ale corneei) care nu au o densitate mare de mitocondrii.



Nimeni nu a comentat acest articol încă.