ARN sau ARN (acid ribonucleic) este un tip de acid nucleic prezent în organismele eucariote și procariote și în viruși. Este un polimer nucleotidic care conține patru tipuri de baze azotate în structura sa: adenină, guanină, citozină și uracil.
ARN-ul se găsește, în general, ca o singură bandă (cu excepția unor viruși), într-un mod liniar sau într-o serie de structuri complexe. De fapt, ARN-ul are un dinamism structural care nu este observat în dubla helix a ADN-ului. Diferitele tipuri de ARN au funcții foarte variate.
ARN-urile ribozomale fac parte din ribozomi, structurile responsabile de sinteza proteinelor în celule. ARN-urile mesager funcționează ca intermediari și transportă informații genetice către ribozom, ceea ce traduce mesajul dintr-o secvență de nucleotide într-o secvență de aminoacizi..
ARN-urile de transfer sunt responsabile pentru activarea și transferul diferitelor tipuri de aminoacizi -20 în total- către ribozomi. Există o moleculă de ARN de transfer pentru fiecare aminoacid care recunoaște secvența din ARN mesager.
În plus, există și alte tipuri de ARN care nu sunt direct implicate în sinteza proteinelor și participă la reglarea genelor..
Indice articol
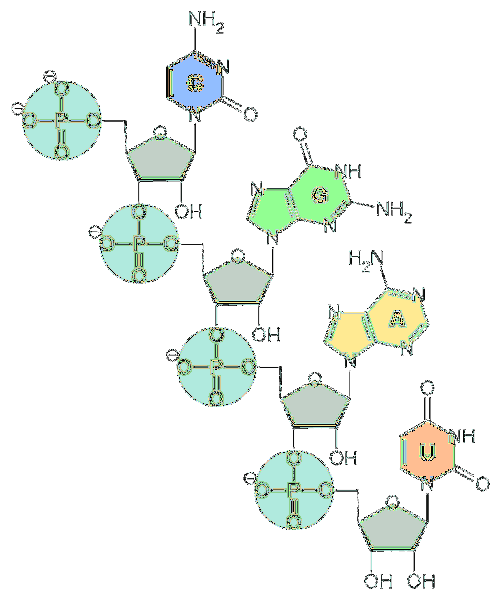
Unitățile fundamentale ale ARN sunt nucleotidele. Fiecare nucleotidă este alcătuită dintr-o bază azotată (adenină, guanină, citozină și uracil), o pentoză și o grupare fosfat..
Bazele de azot sunt derivate din doi compuși fundamentali: pirimidine și purine..
Bazele derivate din purine sunt adenina și guanina, iar bazele derivate din pirimidine sunt citozina și uracilul. Deși acestea sunt cele mai comune baze, acizii nucleici pot avea și alte tipuri de baze mai puțin frecvente..
În ceea ce privește pentozele, acestea sunt unități de d-riboză. Din acest motiv, nucleotidele care alcătuiesc ARN sunt numite „ribonucleotide”.
Nucleotidele sunt legate între ele prin legături chimice care implică gruparea fosfat. Pentru a le forma, gruparea fosfat la capătul 5 'al unei nucleotide este atașată la gruparea hidroxil (-OH) la capătul 3' al următoarei nucleotide, creând astfel o legătură asemănătoare fosfodiesterului..
De-a lungul lanțului de acid nucleic, legăturile fosfodiesterice au aceeași orientare. Prin urmare, există o polaritate a firului, distingând între capătul 3 'și capătul 5'.
Prin convenție, structura acizilor nucleici este reprezentată de capătul 5 'din stânga și capătul 3' din dreapta.
Produsul ARN al transcrierii ADN-ului este o bandă monocatenară care se rotește spre dreapta, într-o conformație elicoidală prin stivuirea bazelor. Interacțiunea dintre purine este mult mai mare decât interacțiunea dintre două pirimidine, datorită dimensiunii lor.
În ARN, nu se poate vorbi de o structură secundară tradițională și de referință, cum ar fi dubla helix a ADN-ului. Structura tridimensională a fiecărei molecule de ARN este unică și complexă, comparabilă cu cea a proteinelor (logic, nu putem globaliza structura proteinelor).
Există interacțiuni slabe care contribuie la stabilizarea ARN-ului, în special stivuirea bazelor, unde inelele sunt situate unul peste altul. Acest fenomen contribuie, de asemenea, la stabilitatea helixului ADN..
Dacă molecula de ARN găsește o secvență complementară, acestea vor fi capabile să cupleze și să formeze o structură cu dublă catenă care se întoarce spre dreapta. Forma predominantă este de tip A; În ceea ce privește formele Z, acestea au fost evidențiate doar în laborator, în timp ce forma B nu a fost observată.
În general, există secvențe scurte (cum ar fi UUGG) care sunt situate la capătul ARN și au particularitatea de a forma bucle grajd. Această secvență participă la plierea structurii tridimensionale a ARN-ului.
În plus, legăturile de hidrogen se pot forma în alte locuri decât perechile tipice de baze (AU și CG). Una dintre aceste interacțiuni are loc între 2'-OH de riboză cu alte grupuri.
Elucidarea diferitelor structuri găsite în ARN a servit pentru a demonstra funcțiile multiple ale acestui acid nucleic..
Există două clase de ARN: informațional și funcțional. Primul grup include ARN-uri care participă la sinteza proteinelor și funcționează ca intermediari în proces; ARN-urile informaționale sunt ARN-uri mesager.
În schimb, ARN-urile aparținând clasei a doua, cele funcționale, nu dau naștere unei noi molecule de proteine, iar ARN-ul în sine este produsul final. Acestea sunt ARN-uri de transfer și ARN-uri ribozomale..
În celulele de mamifere, 80% din ARN este ARN ribozomal, 15% este ARN de transfer și doar o mică porțiune corespunde ARN mesager. Aceste trei tipuri lucrează în cooperare pentru a realiza biosinteza proteinelor..
Există, de asemenea, ARN-uri nucleare mici, ARN-uri citoplasmatice mici și microARN-uri, printre altele. Fiecare dintre cele mai importante tipuri va fi descris în detaliu mai jos:
În eucariote, ADN-ul este limitat la nucleu, în timp ce sinteza proteinelor are loc în citoplasma celulei, unde se găsesc ribozomi. Datorită acestei separări spațiale, trebuie să existe un mediator care să poarte mesajul de la nucleu la citoplasmă și că molecula este ARN-ul mesager..
ARN Messenger, prescurtat mARN, este o moleculă intermediară care conține informațiile codificate în ADN și care specifică o secvență de aminoacizi care va da naștere unei proteine funcționale..
Termenul ARN mesager a fost propus în 1961 de François Jacob și Jacques Monod pentru a descrie porțiunea de ARN care a transmis mesajul de la ADN la ribozomi..
Procesul de sinteză a unui ARNm din catena ADN este cunoscut sub numele de transcripție și are loc diferențial între procariote și eucariote..
Expresia genică este guvernată de mai mulți factori și depinde de nevoile fiecărei celule. Transcrierea este împărțită în trei etape: inițiere, alungire și terminare..
Procesul de replicare a ADN-ului, care are loc în fiecare diviziune celulară, copiază întregul cromozom. Cu toate acestea, procesul de transcriere este mult mai selectiv, se ocupă doar de procesarea unor segmente specifice ale catenei ADN și nu necesită un primer..
Pe Escherichia coli -cea mai bine studiată bacterie din științele vieții - transcrierea începe cu desfacerea dublei spirale ADN și se formează bucla de transcripție. Enzima ARN polimerază este responsabilă pentru sintetizarea ARN și, pe măsură ce transcripția continuă, firul ADN revine la forma sa originală.
Transcrierea nu este inițiată la locații aleatorii pe molecula ADN; există site-uri specializate pentru acest fenomen, numite promotori. Pe E coli ARN polimeraza este cuplată la câteva perechi de baze deasupra regiunii țintă.
Secvențele în care factorii de transcripție sunt cuplați sunt destul de conservați între diferite specii. Una dintre cele mai cunoscute secvențe de promotor este caseta TATA.
La alungire, enzima ARN polimerază adaugă noi nucleotide la capătul 3'-OH, urmând direcția 5 'la 3'. Gruparea hidroxil acționează ca un nucleofil, atacând alfa-fosfatul nucleotidului de adăugat. Această reacție eliberează un pirofosfat.
Doar una dintre catenele de ADN este utilizată pentru a sintetiza ARN-ul mesager, care este copiat în direcția 3 'până la 5' (forma antiparalelă a noii catenă de ARN). Nucleotida care trebuie adăugată trebuie să fie conformă cu împerecherea bazelor: U se împerechează cu A și G cu C.
ARN polimeraza oprește procesul atunci când găsește regiuni bogate în citozină și guanină. În cele din urmă, noua moleculă de ARN mesager este separată de complex.
În procariote, o moleculă de ARN mesager poate codifica mai mult de o proteină.
Atunci când un ARNm codifică exclusiv o proteină sau polipeptidă, se numește ARNm monocistronic, dar dacă codifică mai mult de un produs proteic, ARNm este policistronic (rețineți că în acest context termenul cistron se referă la genă).
În organismele eucariote, marea majoritate a ARNm-urilor sunt monocistronice, iar mașinile de transcripție sunt mult mai complexe în această linie de organisme. Acestea se caracterizează prin faptul că au trei ARN polimeraze, notate I, II și III, fiecare cu funcții specifice.
I este responsabil de sintetizarea pre-ARNr, II sintetizează ARN mesager și unele ARN speciale. În cele din urmă, III este responsabil pentru transferul de ARN-uri, 5S ribozomale și alte ARN-uri mici..
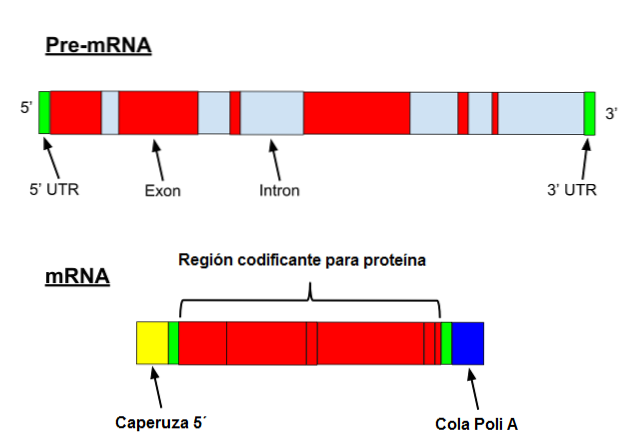
ARN Messenger suferă o serie de modificări specifice în eucariote. Primul presupune adăugarea unui „capac” la capătul 5 '. Din punct de vedere chimic, capacul este un rest de 7-metilguanozină atașat la capăt printr-o legătură 5 ', 5'-trifosfat..
Funcția acestei zone este de a proteja ARN-ul de o posibilă degradare de către ribonucleaze (enzime care descompun ARN-ul în componente mai mici).
În plus, are loc îndepărtarea capătului 3 'și se adaugă 80 până la 250 de reziduuri de adenină. Această structură este cunoscută sub numele de "coadă" poliA și servește ca loc de legare pentru diverse proteine. Când un procariot capătă o coadă poliA, tinde să-i stimuleze degradarea..
Pe de altă parte, acest mesager este transcris cu introni. Intronii sunt secvențe de ADN care nu fac parte din genă, dar care „întrerup” acea secvență. Intronurile nu sunt traduse și, prin urmare, trebuie eliminate din mesager.
Majoritatea genelor vertebratelor au introni, cu excepția genelor care codifică histonele. În mod similar, numărul intronilor dintr-o genă poate varia de la câțiva la zeci dintre aceștia..
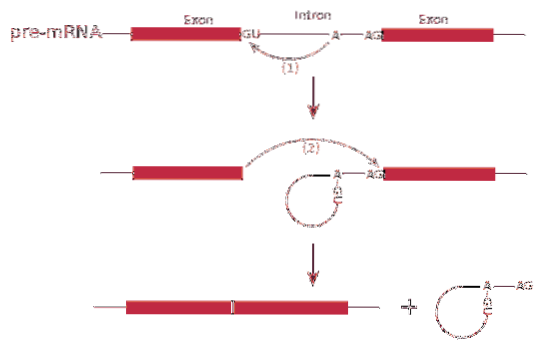
Splicarea ARN sau procesul de îmbinare implică îndepărtarea intronilor din ARN messenger.
Unii introni găsiți în genele nucleare sau mitocondriale pot efectua procesul de îmbinarea fără ajutorul enzimelor sau al ATP. În schimb, procesul se desfășoară prin reacții de transesterificare. Acest mecanism a fost descoperit în protozoanele ciliate Tetrahymena thermophila.
În contrast, există un alt grup de mesageri care nu sunt capabili să medieze pe ai lor îmbinarea, deci au nevoie de utilaje suplimentare. Un număr destul de mare de gene nucleare aparține acestui grup..
Procesul de îmbinarea este mediat de un complex proteic numit spliceozom sau complex de splicing. Sistemul este alcătuit din complexe de ARN specializate numite ribonucleoproteine nucleare mici (RNP)..
Există cinci tipuri de RNP: U1, U2, U4, U5 și U6, care se găsesc în nucleu și mediază procesul de îmbinarea.
îmbinarea poate produce mai multe tipuri de proteine - aceasta este cunoscută sub numele de îmbinarea alternativă, deoarece exonii sunt aranjați diferențiat, creând varietăți de ARN mesager.
ARN ribozomal, prescurtat ARNr, se găsește în ribozomi și participă la biosinteza proteinelor. Prin urmare, este o componentă esențială a tuturor celulelor.
ARN ribozomal se asociază cu molecule proteice (aproximativ 100) pentru a da naștere la presubunități ribozomale. Acestea sunt clasificate în funcție de coeficientul lor de sedimentare, notat cu litera S pentru unitățile Svedberg..
Un ribozom este alcătuit din două părți: subunitatea majoră și subunitatea minoră. Ambele subunități diferă între procariote și eucariote în ceea ce privește coeficientul de sedimentare..
Procariotele au o subunitate mare 50S și o subunitate mică 30S, în timp ce în eucariote subunitatea mare este 60S și cea mică 40S.
Genele care codifică ARN-urile ribozomale se află în nucleol, o anumită zonă a nucleului care nu este delimitată de o membrană. ARN-urile ribozomale sunt transcrise în această regiune de ARN polimeraza I.
În celulele care sintetizează cantități mari de proteine; nucleolul este o structură proeminentă. Cu toate acestea, atunci când celula în cauză nu necesită un număr mare de produse proteice, nucleolul este o structură aproape imperceptibilă.
Subunitatea ribosomală mare 60S este asociată cu fragmentele 28S și 5.8S. În ceea ce privește subunitatea mică (40S), aceasta este asociată cu 18S.
În eucariotele superioare, pre-ARNr este codificat într-o unitate transcripțională 45S, care implică ARN polimeraza I. Acest transcript este procesat în ARN ribosomal 28S, 18S și 5.8S maturi..
Pe măsură ce sinteza continuă, pre-ARNr se asociază cu diferite proteine și formează particule de ribonucleoproteină. Acesta suferă o serie de modificări ulterioare care includ metilarea grupării 2'-OH a ribozei și conversia reziduurilor de uridină în pseudouridină..
Regiunea în care vor apărea aceste modificări este controlată de mai mult de 150 de molecule mici de ARN nucleolar, care au capacitatea de a se cupla cu pre-ARNr..
Spre deosebire de restul pre-ARNr, 5S este transcris de ARN polimeraza III în nucleoplasmă și nu în interiorul nucleolului. După sintetizare, este dus la nucleol pentru a se asambla cu 28S și 5.8S, formând astfel unitățile ribozomale.
La sfârșitul procesului de asamblare, subunitățile sunt transferate în citoplasmă prin porii nucleari..
Se poate întâmpla ca o moleculă de ARN mesager să dea naștere mai multor proteine în același timp, legându-se la mai mult de un ribozom. Pe măsură ce procesul de traducere progresează, sfârșitul mesagerului devine liber și poate fi preluat de un alt ribozom, începând o nouă sinteză..
Din acest motiv, este comun să se găsească ribozomi grupați (între 3 și 10) într-o singură moleculă de ARN mesager, iar acest grup este numit poliribozom.
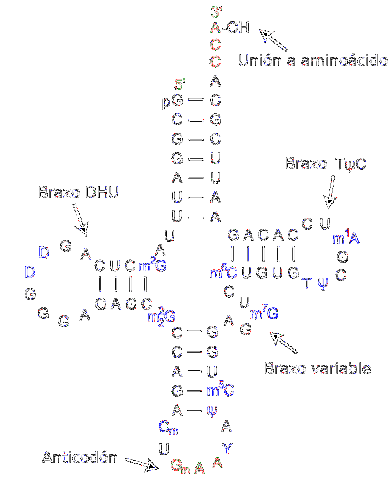
ARN-ul de transfer este responsabil pentru transferul aminoacizilor pe măsură ce progresează procesul de sinteză a proteinelor. Acestea sunt alcătuite din aproximativ 80 de nucleotide (comparativ cu ARN-ul mesager, este o moleculă „mică”).
Structura are falduri și cruci care amintesc de un trifoi cu trei brațe. Un inel adenilic este situat la unul dintre capete, unde gruparea hidroxil a ribozei mediază unirea cu aminoacidul care urmează să fie transportat.
Diferitele ARN-uri de transfer se combină exclusiv cu unul dintre cei douăzeci de aminoacizi care alcătuiesc proteinele; cu alte cuvinte, vehiculul este cel care transportă blocurile de proteine. Complexul ARN de transfer împreună cu aminoacidul se numește aminoacil-ARNt.
Mai mult, în procesul de traducere - care are loc datorită ribozomilor - fiecare ARN de transfer recunoaște un codon specific în ARN-ul mesager. Când este recunoscut, aminoacidul corespunzător este eliberat și devine parte a peptidei sintetizate.
Pentru a recunoaște tipul de aminoacizi care trebuie livrat, ARN-ul are un „anticodon” situat în regiunea de mijloc a moleculei. Acest anticodon este capabil să formeze legături de hidrogen cu bazele complementare prezente în ADN-ul mesager..
MicroARN-urile sau miARN-urile sunt un tip de ARN monocatenar scurt, între 21 și 23 de nucleotide, a cărui funcție este de a regla expresia genelor. Deoarece nu este tradus în proteine, este adesea numit ARN necodificator..
La fel ca alte tipuri de ARN, procesarea microARN este complexă și implică o serie de proteine.
MicroARN-urile apar din precursori mai lungi numiți mi-pri-ARN-uri, derivate din primul transcript al genei. În nucleul celulei, acești precursori sunt modificați în complexul microprocesor și rezultatul este un pre-miARN..
Pre-miARN-urile sunt 70 de agrafe de nucleotide care își continuă procesarea în citoplasmă de către o enzimă numită Dicer, care asamblează complexul de tăcere indus de ARN (RISC) și în final miARN-ul este sintetizat..
Acești ARN sunt capabili să regleze expresia genelor, deoarece sunt complementare ARN-urilor mesagere specifice. Prin împerecherea cu ținta lor, miARN-urile sunt capabile să suprime mesagerul sau chiar să-l degradeze. În consecință, ribozomul nu poate traduce transcrierea respectivă..
Un tip particular de microARN este ARN-urile mici care interferează (ARNsi), numite și ARN-uri de tăcere. Sunt ARN-uri scurte, între 20 și 25 de nucleotide, care împiedică exprimarea anumitor gene.
Sunt instrumente foarte promițătoare pentru cercetare, deoarece fac posibilă reducerea la tăcere a unei gene de interes și astfel studierea funcției sale posibile..
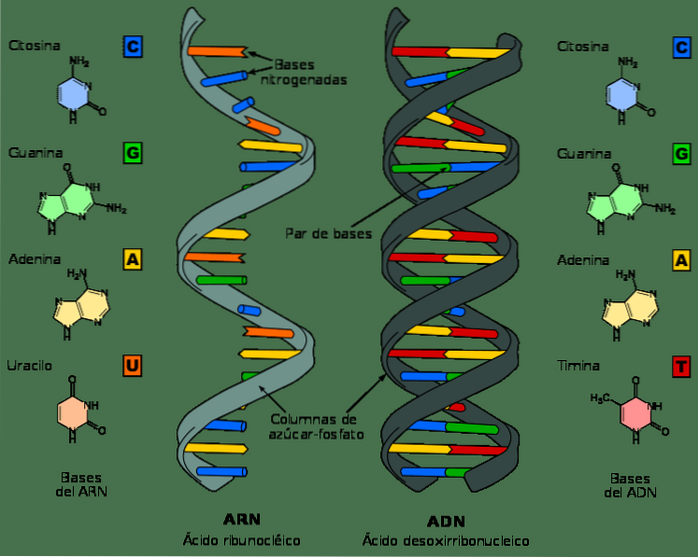
Deși ADN-ul și ARN-ul sunt acizi nucleici și pot arăta foarte asemănător la prima vedere, ele diferă prin mai multe dintre proprietățile lor chimice și structurale. ADN-ul este o moleculă cu bandă dublă, în timp ce ARN-ul este cu o singură bandă.
Ca urmare, ARN-ul este o moleculă mai versatilă și poate lua o mare varietate de forme tridimensionale. Cu toate acestea, anumiți viruși au ARN cu bandă dublă în materialul lor genetic..
În nucleotidele de ARN, molecula de zahăr este o riboză, în timp ce în ADN este o dezoxiriboză, diferind doar în prezența unui atom de oxigen.
Legătura fosfodiesterică din coloana vertebrală a ADN-ului și ARN-ului este predispusă la un proces lent de hidroliză fără prezența enzimelor. În condiții alcaline, ARN-ul se hidrolizează rapid - datorită grupului hidroxil suplimentar - în timp ce ADN-ul nu.
În mod similar, bazele azotate care alcătuiesc nucleotidele din ADN sunt guanina, adenina, timina și citozina; în contrast, în ARN, timina este înlocuită cu uracil. Uracil se poate împerechea cu adenină, în același mod ca timina din ADN.
ARN este singura moleculă cunoscută capabilă să stocheze informații și să catalizeze reacții chimice în același timp; Din acest motiv, mai mulți autori propun că molecula de ARN a fost crucială la originea vieții. În mod surprinzător, substraturile ribozomilor sunt alte molecule de ARN.
Descoperirea ribozimelor a dus la redefinirea biochimică a "enzimei" - deoarece anterior termenul era folosit exclusiv pentru proteinele cu activitate catalitică - și a ajutat la susținerea unui scenariu în care primele forme de viață foloseau doar ARN ca material genetic.



Nimeni nu a comentat acest articol încă.