microfilamente sau filamente de actină, sunt una dintre cele trei componente fundamentale ale citoscheletului celulelor eucariote (microfilamente, microtubuli și filamente intermediare) și sunt compuse din filamente mici ale unei proteine numite actină (polimeri de actină).
În eucariote, genele care codifică microfilamentele de actină sunt foarte conservate în toate organismele, motiv pentru care sunt adesea utilizate ca markeri moleculari pentru studii de diferite tipuri.

Microfilamentele sunt distribuite în întregul citosol, dar sunt deosebit de abundente în regiunea care stă la baza membranei plasmatice, unde formează o rețea complexă și se asociază cu alte proteine speciale pentru a forma citoscheletul..
Rețelele de microfilamente din citoplasma celulelor de mamifere sunt codificate de două dintre cele șase gene descrise pentru actină, care sunt implicate în dinamica microfilamentelor și sunt chiar foarte importante în timpul diferențierii celulelor stem.
Mulți autori sunt de acord că microfilamentele sunt cele mai diverse, versatile și importante proteine din citoscheletul majorității celulelor eucariote și este important să ne amintim că acestea nu se găsesc în microorganismele procariote..
Pe acest tip de celule, pe de altă parte, există filamente care sunt omoloage microfilamentelor, dar care sunt alcătuite dintr-o altă proteină: proteina MreB..
În prezent, se crede că gena care codifică această proteină este o posibilă genă ancestrală pentru actina eucariotă. Cu toate acestea, omologia secvenței aminoacizilor care alcătuiesc proteina MreB este de numai 15% față de secvența actinică.
Deoarece sunt o parte fundamentală a citoscheletului, orice defect fenotipic atât în microtubuli, cât și în filamente intermediare și microfilamente de actină (citoschelet) poate provoca diferite patologii celulare și sistemice.
Indice articol
Microfilamentele sunt alcătuite din proteine monomeri din familia actinelor, care sunt proteine contractile foarte abundente în celulele eucariote, deoarece participă și la contracția musculară..
Aceste filamente au un diametru cuprins între 5 și 7 nm, deci sunt cunoscute și sub numele de filamente subțiri și sunt compuse din două forme de actină: forma globulară (actina G) și forma filamentoasă (actina F)..
Proteinele care participă la citoschelet sunt cunoscute sub numele de actine γ și β, în timp ce cele care participă la contracție sunt de obicei actine α..
Proporția de actină globulară și actină filamentoasă din citosol depinde de nevoile celulare, deoarece microfilamentele sunt structuri foarte variabile și versatile, care sunt în continuă creștere și se scurtează prin polimerizare și depolimerizare..
Actina G este o proteină globulară mică, formată din aproape 400 de aminoacizi și cu o greutate moleculară de aproximativ 43 kDa.
Monomerii G-actină care alcătuiesc microfilamentele sunt aranjați sub forma unei suvite elicoidale, deoarece fiecare suferă o răsucire atunci când este asociată cu următoarea.
Actina G se asociază cu o moleculă de Ca2 + și o altă ATP, care îi stabilizează forma globulară; în timp ce F-actina se obține după hidroliza fosfatului terminal al moleculei de ATP în G-actină, ceea ce contribuie la polimerizare.
Filamentele de actină pot fi organizate în „pachete” sau „rețele” care au funcții diferite în interiorul celulelor. Fasciculele formează structuri paralele legate de punți transversale destul de rigide.
Rețelele, pe de altă parte, sunt structuri mai libere, precum ochiurile tridimensionale cu proprietățile gelurilor semi-solide..
Există multe proteine care sunt asociate cu filamente de actină sau microfilamente și sunt cunoscute sub numele de ABP (din engleză proteine care leagă actina), care au site-uri specifice pentru aceasta.
Multe dintre aceste proteine permit microfilamentelor să interacționeze cu celelalte două componente ale citoscheletului: microtubuli și filamente intermediare, precum și cu celelalte componente ale feței interioare a membranei plasmatice..
Alte proteine cu care interacționează microfilamentele includ laminele nucleare și spectrina (în celulele roșii din sânge).
Deoarece monomerii actinici globulari se leagă întotdeauna în același mod, orientați în aceeași direcție, microfilamentele au o polaritate definită, cu două capete: unul „mai mult” și unul „mai puțin”..
Polaritatea acestor filamente este foarte importantă, deoarece acestea cresc considerabil mai repede la capătul lor pozitiv, unde sunt adăugați noii monomeri G-actină..

Primul lucru care are loc în timpul polimerizării filamentelor de actină este un proces cunoscut sub numele de „nucleație”, care constă în asocierea a trei monomeri ai proteinei.
La acest trimer se adaugă monomeri noi la ambele capete, astfel încât filamentul să crească. Monomerii G-actină sunt capabili să hidrolizeze ATP cu fiecare legare, ceea ce are implicații asupra ratei de polimerizare, deoarece fragmentele actină-ATP se disociază cu o dificultate mai mare decât fragmentele actină-ADP..
ATP nu este necesar pentru polimerizare și rolul specific al hidrolizei sale nu a fost încă elucidat..
Unii autori consideră că, deoarece evenimentele de polimerizare a actinei sunt reversibile rapid, ATP asociat cu aceste procese poate reprezenta până la 40% din cifra de afaceri totală celulară a acestei molecule de energie.
Atât polimerizarea filamentelor de actină, cât și depolimerizarea lor sunt procese extrem de reglementate de o serie de proteine specifice, care sunt responsabile pentru remodelarea filamentelor..
Un exemplu de proteine care reglează depolimerizarea sunt factorul de depolimerizare a actinei cofilină. O altă proteină, profilina, are o funcție opusă, deoarece stimulează asocierea monomerilor (prin stimularea schimbului de ADP cu ATP).
Microfilamentele interacționează cu filamentele de miozină care sunt asociate cu proteinele transmembranare care au un domeniu în citosol și un altul în exteriorul celulei, participând astfel la procesele de mobilitate celulară.
Aceste microfilamente asociate cu membrana plasmatică mediază diferite răspunsuri celulare la diferite clase de stimuli. De exemplu, adeziunea celulară în țesuturile epiteliale este condusă de proteinele transmembranare cunoscute sub numele de cadherine, care interacționează cu microfilamentele pentru a recruta factorii de răspuns..
Filamentele de actină interacționează cu filamentele intermediare pentru a determina transmiterea stimulilor extracelulari către locații cheie, cum ar fi ribozomii și cromozomii din nucleu..
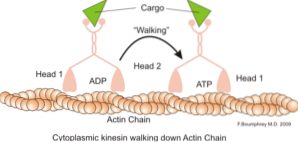
O funcție clasică și mult studiată a microfilamentelor este capacitatea lor de a forma "punți", "șine" sau "autostrăzi" pentru mișcarea proteinei miozina I, care este capabilă să încarce vezicule de transport de la organite la membrana plasmatică în secretor căi.
Microfilamentele interacționează, de asemenea, cu mioza II pentru a stabili inelul contractil care se formează în timpul citokinezei, tocmai în timpul ultimei etape a diviziunii celulare în care citosolul este separat de celulele stem și celule fiice..
În general, microfilamentele cu F-actină modulează distribuția unor organite, cum ar fi complexul Golgi, reticulul endoplasmatic și mitocondriile. În plus, ei participă, de asemenea, la poziționarea spațială a ARNm, astfel încât acestea să fie citite de ribozomi..
Întregul set celular de microfilamente, în special cele care sunt strâns legate de membrana plasmatică, participă la formarea membranelor ondulate ale celulelor care au o mișcare activă constantă.
Ele sunt, de asemenea, implicate în formarea de microvili și alte umflături comune la suprafața multor celule..
Microfilamentele participă la procesul de secreție biliară în hepatocite (celule hepatice) și, de asemenea, la mișcările peristaltice (contracție coordonată) ale canaliculilor hepatici.
Acestea contribuie la diferențierea domeniilor membranei plasmatice datorită asocierii lor cu diferite elemente citosolice și controlului pe care îl exercită asupra topografiei acestor elemente intracelulare..
Există puține boli asociate cu defecte primare în structură sau cu proteine și enzime reglatoare în sinteza microfilamentelor, în ciuda faptului că acestea sunt direct implicate într-un număr mare de funcții..
Rata scăzută de boli și malformații în structura primară a microfilamentelor se datorează faptului că, în general, există gene multiple care codifică atât actina, cât și proteinele sale reglatoare, fenomen cunoscut sub numele de „redundanță genetică”..
Una dintre cele mai studiate patologii este vitrificarea ovocitelor pe citoscheletul lor, unde se observă o întrerupere a rețelei de microfilamente corticale, precum și o depolimerizare și dezorganizare a microtubulilor fusului mitotic..
În termeni generali, această vitrificare determină o dispersie cromozomială, deoarece duce la o tulburare a compactării întregii cromatine.
Celulele care au o organizare și o proporție mai mare de microfilamente în citoscheletul lor sunt celulele mușchiului striat, prin urmare, majoritatea patologiilor sunt asociate cu o defecțiune a aparatului contractil..
Microfilamentele defecte sau atipice au fost, de asemenea, asociate cu boala osoasă cunoscută sub numele de boala Paget..


Nimeni nu a comentat acest articol încă.