citocrom c oxidază Este un complex de proteine enzimatice care poate traversa bistratul lipidic al membranei celulare. Este hemosolubil și este în principal asociat cu membrana interioară a mitocondriilor, fiind găsit atât în organismele procariote (bacterii), cât și în eucariote (unicelulare și multicelulare)..
De asemenea, numită complex IV, această enzimă este vitală în funcțiile metabolice aerobe ale organismelor, deoarece este esențială în lanțul de transport al electronilor în care celula arde zaharuri și captează o parte din energia eliberată pentru a stoca adenozin trifosfat sau ATP..
Denumirea de citocrom înseamnă „pigmenți celulari”. Acestea sunt proteine din sânge care transportă electroni. Citocromii au fost descoperiți de medicul irlandez Charles Alexander MacMunn în 1884. MacMunn a fost pionierul descoperirii pigmentului respirator din sânge, numit astăzi citocrom 1..
În anii 1920, entomologul și parazitologul rus David Keilin a redescoperit și caracterizat pigmenții respiratori și a fost cel care le-a numit citocromi. Deși MacMunn le descoperise în 1884, comunitatea științifică îl uitase și unii chiar i-au interpretat greșit opera..
Indice articol
În general, pigmenții respiratori au spectre caracteristice ale luminii vizibile. Se știe că există cel puțin patru complexe proteice de membrană integrate în care există 5 tipuri diferite de citocromi: a, a3, b, c1 și c, clasificate în funcție de lungimile de undă ale absorbției spectrale maxime.
Ele se găsesc în general pe membrana interioară a mitocondriilor. Cu toate acestea, au fost observate și în reticulul endoplasmatic și în cloroplaste, în organismele eucariote și procariote..
Prezintă grupul protetic hemic conținând fier (Fe). Fiecare dintre citocromii cunoscuți acționează în complexe multienzimale în transportul electronilor în procesul respirator sau în lanț..
Citocromii au funcția de a participa la reacțiile de oxidare-reducere. Reacțiile de reducere, unde acceptă electroni, apar diferit pentru fiecare tip de citocrom, iar valoarea lor este determinată de fluxul de electroni din lanțul respirator.
Se cunosc patru clase de citocrom c, care sunt după cum urmează.
În cadrul acestei clase se află citocromii solubili cu spin redus c (lowspin), prezenți în bacterii și mitocondrii. Sunt octaedrici. Situsul de legare a hemului este la capătul N-terminal al histidinei, iar al șaselea ligand este furnizat de un reziduu de metionină la capătul C-terminal..
Din această clasă, pot fi recunoscute chiar mai multe subclase, a căror structură tridimensională a fost determinată..
Citocromul cu spin mare și unele cu citocromul cu spin mic se găsesc în această clasă. Cele cu rotație înaltă au situsul de legare aproape de capătul C-terminal, iar în cele cu rotație mică al șaselea ligand pare a fi un reziduu de metionină lângă capătul N-terminal. Acestea sunt pentacoordonate cu histidina celui de-al cincilea ligand.
Această clasă se caracterizează prin prezentarea citocromilor c cu hem multiplu (c3 și c7) și un potențial redus de reducere a oxidului, cu doar 30 de reziduuri de aminoacizi pe grupă hem. La reprezentanții acestei clase, grupurile hem c au structură și funcții neechivalente, pe lângă faptul că prezintă potențiale redox diferite. Sunt octaedrici.
Potrivit unor autori, această clasă a fost creată numai pentru a include proteine complexe pe care le prezintă alte grupuri protetice, precum și hemul c sau flavocitocromul c, printre altele..
Citocrom c oxidaza este o enzimă mitocondrială care efectuează faza finală a transportului de electroni în respirația celulară. Această enzimă catalizează transportul electronilor din citocromul redus c în oxigen..
Unii compuși chimici, cum ar fi cianura, dioxidul de carbon și azida, pot inhiba funcționarea acestei enzime, provocând așa-numita asfixie chimică celulară. Alte forme de inhibare a complexului IV sunt mutațiile genetice.
Din punct de vedere evolutiv, citocrom c oxidaza se găsește numai în organismele aerobe și mai multe grupuri de oameni de știință sugerează că prezența acestei proteine indică relații evolutive în care plantele, ciupercile și animalele au împărtășit un strămoș comun..
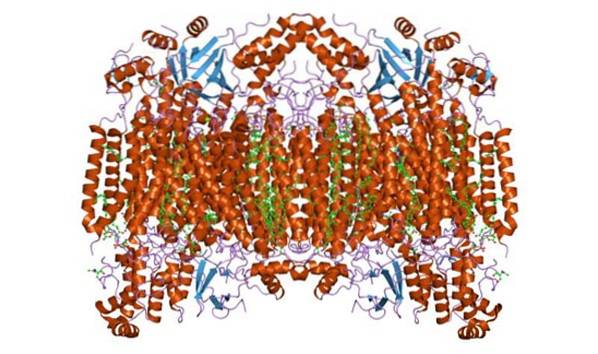
Citocrom c oxidaza formează un complex homodimeric, adică compus din doi monomeri similari în membrana interioară a mitocondriilor. Complexul enzimatic este format din 3 până la 4 subunități în organismele procariote și până la maximum 13 (unii sugerează 14) polipeptide în organisme precum mamiferele..
La aceste organisme, 3 polipeptide sunt de origine mitocondrială, iar restul provin din nucleu. Fiecare monomer are 28 de spirale transmembranare care separă domeniile hidrofile de matricea membranei și spațiul intermembranar..
Are o singură unitate catalitică, care se găsește în toate enzimele care catalizează reacțiile de oxidare / reducere, folosind oxigen molecular (oxidaze, în special hem-cupru). Complexul conține citocromi a și a3 legați de subunitatea I și doi centri de cupru.
Are una sau mai multe grupe hemo C legate de structura proteică înconjurătoare prin una sau mai multe (în general două) legături tioeter. Alți autori sugerează că există o singură grupă hemo C legată covalent de proteină între inelul porfirină și două reziduuri de cisteină..
Singura grupă hemo c menționată mai sus este înconjurată de reziduuri hidrofobe și este hexacoordonată, cu histidină în poziția 18 a lanțului polipeptidic și metionină în poziția 80..
Citoxromul c oxidazele sunt protagoniști în trei mecanisme fiziologice principale, pe care le vom vedea mai jos.
Apoptoza este distrugerea celulară sau moartea programată, cauzată de organismul însuși și al cărei scop este de a controla creșterea, dezvoltarea, eliminarea țesuturilor deteriorate și reglarea sistemului imunitar. În acest proces fiziologic, citocrom c oxidaza participă ca intermediar.
Această proteină, eliberată de mitocondrii, duce la o interacțiune cu reticulul endoplasmatic, care determină secreția sau eliberarea de calciu. Creșterea progresivă a calciului declanșează o eliberare masivă de citocrom c oxidază, până când sunt atinse nivelurile citotoxice de calciu..
Nivelurile citotoxice de calciu și eliberarea citocromelor c determină activarea în cascadă a mai multor enzime caspazice, care sunt responsabile pentru distrugerea celulelor.
Mai multe studii indică faptul că atunci când citocrom c oxidaza este expusă la lungimi de undă de 670 nanometri, aceasta participă la un complex funcțional care pătrunde în țesutul deteriorat sau rănit și crește rata de regenerare celulară..
Aceasta este probabil cea mai cunoscută și cea mai relevantă funcție a citocrom c oxidazei. Tocmai complexul oxidazic (al lanțului respirator) este responsabil pentru colectarea electronilor din citocromul c și transferarea lor către molecula de oxigen, reducându-l la două molecule de apă..
Conectat la acest proces, o translocație de protoni are loc prin membrană, rezultând generarea unui gradient electrochimic pe care complexul ATP sintetază îl folosește pentru a produce sau sintetiza ATP (adenozin trifosfat)..
Citocrom c oxidaza este inhibată de diferiți compuși chimici și procese. Modul în care apare poate apărea ca o modalitate naturală de a regla producția sau acțiunea enzimei sau poate apărea accidental din cauza otrăvirii.
În prezența azidei, cianurii sau monoxidului de carbon, citocrom c oxidaza se leagă de acestea și funcționarea complexului proteic este inhibată. Acest lucru provoacă o întrerupere a procesului respirator celular și, prin urmare, provoacă sufocarea chimică a celulelor..
Alți compuși, cum ar fi oxidul azotic, hidrogenul sulfurat, metanolul și unii alcooli metilați, provoacă, de asemenea, inhibarea citocromului c-oxidazei..
Citocrom c oxidaza este o enzimă care este controlată de gene atât în nucleu, cât și în mitocondrie. Există modificări genetice sau mutații care pot duce la un deficit de citocrom c oxidază.
Aceste mutații perturbă funcționalitatea enzimei, deoarece îi modifică structura enzimatică, aducând cu ea tulburări metabolice în timpul dezvoltării embrionare (conform studiilor efectuate la oameni), care vor afecta ulterior organismul în primii săi ani de viață..
Deficitul de citocrom c oxidază afectează țesuturile cu cerere mare de energie, cum ar fi inima, ficatul, creierul și mușchii. Simptomele acestor mutații se reflectă înainte de cei doi ani de viață și se pot manifesta ca afecțiuni puternice sau ușoare.
Simptomele ușoare pot fi observate chiar la scurt timp după primul an de vârstă, iar indivizii cu acestea au de obicei doar tensiune musculară scăzută (hipotonie) și atrofie musculară (miopatie)..
Pe de altă parte, persoanele cu simptome mai puternice pot avea atrofie musculară și encefalomiopatie. Alte afecțiuni cauzate de absența citocromului c oxidază sunt cardiomiopatia hipertrofică, mărirea patologică a ficatului, sindromul Leigh și acidoză lactică..
Filogenia este știința responsabilă de studiile originii, formării și dezvoltării evolutive din punct de vedere strămoș-descendent, al organismelor. În ultimele decenii, studiile de filogenie cu analiză moleculară au fost din ce în ce mai frecvente, oferind o mulțime de informații și rezolvând probleme taxonomice..
În acest sens, unele studii filogenetice indică faptul că utilizarea citocromului c oxidazelor poate ajuta la stabilirea unor relații evolutive. Acest lucru se datorează faptului că acest complex proteic este foarte conservat și este prezent într-o mare varietate de organisme, variind de la protiști unicelulari până la vertebrate mari..
Un exemplu în acest sens sunt testele făcute cu oameni, cimpanzei (Pâine Paniscus) și macacii Rhesus (Macaca mulatta). Astfel de teste au dezvăluit că moleculele citocrom c oxidazei umane și ale cimpanzeului erau identice..
De asemenea, a arătat că moleculele de citocrom c oxidază ale macacului Rhesus diferă cu un aminoacid de cele din primii doi, reafirmând în consecință relațiile strămoș-descendenți dintre cimpanzei și oameni..



Nimeni nu a comentat acest articol încă.