
sfingolipide ele reprezintă una dintre cele trei mari familii de lipide prezente în membranele biologice. La fel ca glicerofosfolipidele și sterolii, acestea sunt molecule amfipatice cu o regiune polară hidrofilă și o regiune apolară hidrofobă..
Au fost descrise pentru prima dată în 1884 de Johann L. W. Thudichum, care a descris trei sfingolipide (sfingomielină, cerebrozide și cerebrosulfatide) care aparțin celor trei clase diferite cunoscute: fosfosfingolipide, glicozifingolipide neutre și acide..
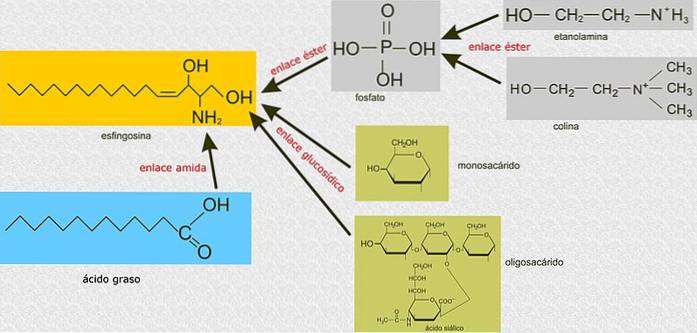
Spre deosebire de glicerofosfolipide, sfingolipidele nu sunt construite pe o moleculă de glicerol 3-fosfat ca schelet principal, ci sunt compuși derivați din sfingozină, un amino alcool cu un lanț lung de hidrocarburi legat printr-o legătură amidică..
În ceea ce privește complexitatea și diversitatea, cel puțin 5 tipuri de baze diferite sunt cunoscute pentru sfingolipide la mamifere. Aceste baze pot fi alăturate de mai mult de 20 de tipuri diferite de acizi grași, cu lungimi și grade variabile de saturație, pe lângă multiplele variații ale grupurilor polare care pot apărea..
Membranele biologice au aproximativ 20% sfingolipide. Acestea au funcții variate și importante în celule, de la transducție structurală la semnal, și controlul diferitelor procese de comunicare celulară..
Distribuția acestor molecule variază în funcție de funcția organului unde se găsesc, dar în mod normal concentrația de sfingolipide este mult mai mare în monostratul exterior al membranei plasmatice în comparație cu monostratul interior și alte compartimente..
La om există cel puțin 60 de specii de sfingolipide. Multe dintre ele sunt componente importante ale membranelor celulelor nervoase, în timp ce altele joacă roluri structurale importante sau participă la transducția semnalului, recunoașterea, diferențierea celulară, patogeneza, moartea celulară programată, printre altele..
Indice articol
Toate sfingolipidele sunt derivate dintr-o L-serină, care este condensată cu un acid gras cu lanț lung pentru a forma baza sfingoidă, cunoscută și sub numele de bază cu lanț lung (LCB)..
Cele mai frecvente baze sunt sfinganina și sfingozina, care diferă între ele numai în prezența unei legături duble trans între carbonii 4 și 5 ai acidului gras al sfingozinei.
Carbonii 1, 2 și 3 ai sfingozinei sunt analogi din punct de vedere structural cu carbonii glicerolului glicerofosfolipidelor. Când un acid gras este atașat la carbonul 2 al sfingozinei prin legături amidice, se produce o ceramidă, care este o moleculă foarte asemănătoare diacilglicerolului și reprezintă cel mai simplu sfingolipid.
Acizii grași cu lanț lung care alcătuiesc regiunile hidrofobe ale acestor lipide pot fi foarte diverse. Lungimile variază de la 14 la 22 de atomi de carbon care pot avea diferite grade de saturație, de obicei între carbonii 4 și 5.
În pozițiile 4 sau 6 pot avea grupări hidroxil și duble legături în alte poziții sau chiar ramuri precum grupările metil.
Lanțurile de acizi grași legați prin legături amidice de ceramide sunt în mod obișnuit saturate și tind să fie mai lungi decât cele găsite în glicerofosfolipide, ceea ce pare a fi crucial pentru activitatea lor biologică..
O trăsătură distinctivă a scheletului sfingolipidic este că pot avea o încărcătură netă pozitivă la pH neutru, rară printre moleculele lipidice..
Însă pKa grupului amino este scăzut în comparație cu o amină simplă, între 7 și 8, astfel încât o porțiune a moleculei nu este încărcată la pH fiziologic, ceea ce ar putea explica mișcarea „liberă” a acestora între straturile bistratice.
Clasificarea tradițională a sfingolipidelor rezultă din multiplele modificări pe care le poate suferi molecula de ceramidă, în special în ceea ce privește substituțiile grupurilor capului polar.
Sfingolipidele sunt esențiale la animale, plante și ciuperci, precum și la unele organisme procariote și viruși..
Sfingolipidele modulează proprietățile fizice ale membranelor, inclusiv fluiditatea, grosimea și curbura acestora. Modularea acestor proprietăți le conferă, de asemenea, influență directă asupra organizării spațiale a proteinelor de membrană..
În membranele biologice, pot fi detectate micro-domenii dinamice cu fluiditate mai mică, alcătuite din molecule de colesterol și sfingolipide numite plute lipidice..
Aceste structuri apar în mod natural și sunt strâns legate de proteinele integrale, receptorii de suprafață celulară și proteinele de semnalizare, transportorii și alte proteine cu ancore glicozilfosfatidilinozitol (GPI)..
Au funcții ca molecule de semnalizare care acționează ca al doilea mesager sau ca liganzi secreti pentru receptorii de pe suprafața celulei..
Ca mesageri secundari, ei pot participa la reglarea homeostaziei calciului, a creșterii celulare, a tumorigenezei și a suprimării apoptozei. Mai mult, activitatea multor proteine de membrană integrală și periferică depinde de asocierea lor cu sfingolipidele..
Multe interacțiuni între celule și celulă cu mediul său depind de expunerea diferitelor grupuri polare de sfingolipide la fața exterioară a membranei plasmatice..
Legarea glicozifingolipidelor și lectinelor este crucială pentru asocierea mielinei cu axoni, aderența neutrofilelor la endoteliu etc..
Cele mai importante sfingolipide de semnalizare sunt bazele cu lanț lung sau sfingozinele și ceramidele, precum și derivații lor fosforilați, cum ar fi sfingozina 1-fosfatul.
Produsele metabolice ale multor sfingolipide activează sau inhibă mai multe ținte din aval (protein kinaze, fosfoproteine fosfataze și altele), care controlează comportamente celulare complexe, cum ar fi creșterea, diferențierea și apoptoza..
Unii agenți patogeni utilizează glicosfingolipidele ca receptori pentru a media intrarea lor în celulele gazdă sau pentru a le furniza factori de virulență..
S-a demonstrat că sfingolipidele participă la mai multe evenimente celulare, cum ar fi secreția, endocitoza, chimiotaxia, neurotransmisia, angiogeneza și inflamația..
Sunt, de asemenea, implicați în traficul de membrane, influențând astfel internalizarea receptorilor, ordonarea, mișcarea și fuziunea veziculelor secretoare ca răspuns la diferiți stimuli..
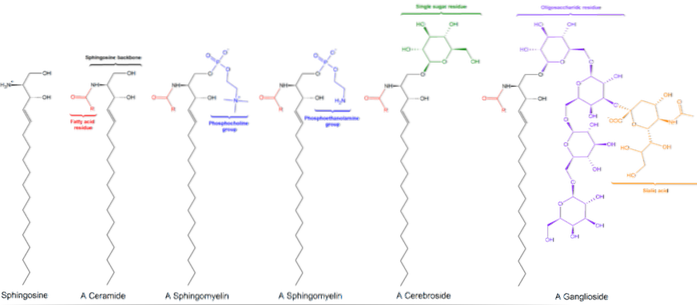
Există trei subclase de sfingolipide, toate derivate din ceramidă și care diferă între ele prin grupe polare, și anume: sfingomieline, glicolipide și gangliozide..

Acestea conțin fosfocolină sau fosfoetanolamină ca grup cap polar, motiv pentru care sunt clasificate ca fosfolipide împreună cu glicerofosfolipide. Ele seamănă, desigur, cu fosfatidilcoline în structură tridimensională și proprietăți generale, deoarece nu au nicio încărcare pe capetele lor polare..
Acestea sunt prezente în membranele plasmatice ale celulelor animale și sunt deosebit de abundente în mielină, o teacă care înconjoară și izolează axonii unor neuroni.
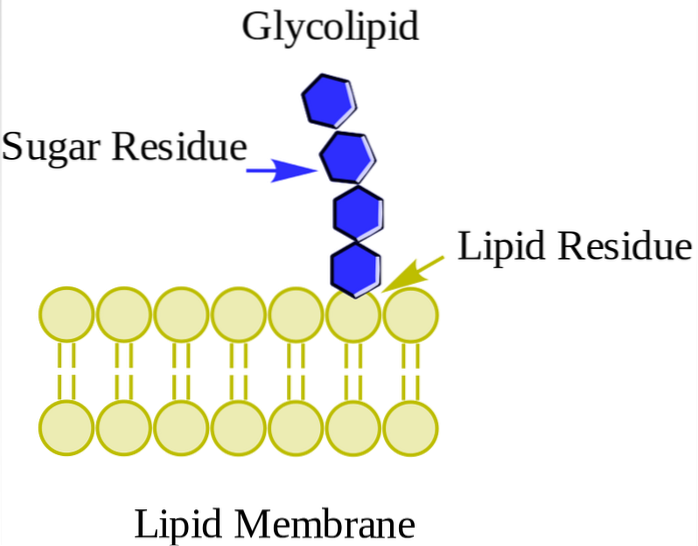
Acestea se găsesc în principal pe fața exterioară a membranei plasmatice și au unul sau mai multe zaharuri sub formă de grup polar polar atașat direct la hidroxilul de carbon 1 al porțiunii de ceramidă. Nu au grupe fosfat. Deoarece la pH 7 nu au nicio încărcare, se numesc glicolipide neutre.
Cerebrozidele au o singură moleculă de zahăr atașată la ceramidă. Cele care conțin galactoză se găsesc în membranele plasmatice ale celulelor țesutului ne-nervos. Globozidele sunt glicozifingolipide cu două sau mai multe zaharuri, de obicei D-glucoză, D-galactoză sau N-acetil-D-galactozamină..
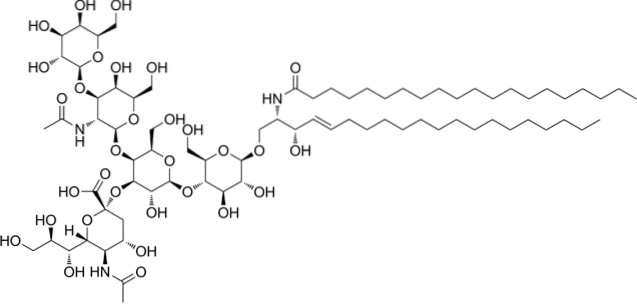
Acestea sunt cele mai complexe sfingolipide. Au oligozaharide ca grup cap polar și unul sau mai multe reziduuri terminale de acid N-acetilmuramic, numite și acid sialic. Acidul sialic conferă gangliozidelor o sarcină negativă la pH 7, ceea ce le distinge de glicozifingolipide neutre.
Nomenclatura acestei clase de sfingolipide depinde de cantitatea de reziduuri de acid sialic prezente în porțiunea oligozaharidică a capului polar..
Molecula de bază cu lanț lung, sau sfingozină, este sintetizată în reticulul endoplasmatic (ER) și adăugarea grupului polar la capul acestor lipide are loc mai târziu în complexul Golgi. La mamifere, o anumită sinteză a sfingolipidelor poate apărea și în mitocondrii.
După finalizarea sintezei lor în complexul Golgi, sfingolipidele sunt transportate către alte compartimente celulare prin mecanisme mediate de vezicule..
Biosinteza sfingolipidelor constă din trei evenimente fundamentale: sinteza bazelor cu lanț lung, biosinteza ceramidelor prin unirea unui acid gras printr-o legătură amidică și, în cele din urmă, formarea sfingolipidelor complexe prin uniunea grupurilor polare pe carbonul 1 al bazei sfingoide.
Pe lângă sinteză de novo, sfingolipidele pot fi, de asemenea, formate prin rotirea sau reciclarea bazelor cu lanț lung și a ceramidelor, care pot alimenta rezerva de sfingolipide.
Biosinteza ceramidei, coloana vertebrală a sfingolipidelor, începe cu condensarea decarboxilativă a unei molecule de palmitoil-CoA și a unei L-serine. Reacția este catalizată de o serin palmitoil transferază heterodimerică (SPT), dependentă de fosfat piridoxal și produsul este 3-ceto dihidrosfingozină..
Această enzimă este inhibată de β-halo-L-alanine și L-cicloserine. În drojdie este codificată de două gene, în timp ce la mamifere există trei gene pentru această enzimă. Situl activ este situat pe fața citoplasmatică a reticulului endoplasmatic.
Rolul acestei prime enzime este conservat la toate organismele studiate. Cu toate acestea, există unele diferențe între taxonii care au legătură cu localizarea subcelulară a enzimei: cea a bacteriilor este citoplasmatică, cea a drojdiilor, plantelor și animalelor se află în reticulul endoplasmatic..
3-ketosfinganina este ulterior redusă de 3-ketosfinganină reductază dependentă de NADPH pentru a produce sfinganină. Dihidroceramidă sintază (sfinganină N-acil transferază) apoi acetilează sfinganină pentru a produce dihidroceramidă. Ceramida este apoi formată din dihidroceramidă desaturază / reductază, care introduce o legătură dublă trans în poziția 4-5.
La mamifere există numeroase izoforme de ceramidă sintază, fiecare legând un lanț specific de acizi grași de bazele cu lanț lung. Prin urmare, ceramidele sintaze și alte enzime, elongazele, reprezintă principala sursă de diversitate pentru acizii grași din sfingolipide..
Sfingomielina este sintetizată prin transferul unei fosfocoline din fosfatidilcolină în ceramidă, eliberând diacilglicerol. Reacția leagă căile de semnalizare sfingolipid și glicerofosfolipid.
Fosfoetanolamina ceramidă este sintetizată din fosfatidiletanolamină și ceramidă într-o reacție similară cu cea a sintezei de sfingomielină și, odată formată, poate fi metilată cu sfingomielină. Ceramidele de inozitol fosfat se formează prin transesterificare din fosfatidilinozitol.
Glicozifingolipidele sunt modificate în principal în complexul Golgi, unde enzimele specifice glicoziltransferazei participă la adăugarea lanțurilor oligozaharidice în regiunea hidrofilă a scheletului ceramidei..
Degradarea sfingolipidelor este realizată de enzimele glucohidrolaze și sfingomielinaze, care sunt însărcinate cu eliminarea modificărilor grupurilor polare. Pe de altă parte, ceramidazele regenerează bazele cu lanț lung din ceramide.
Gangliozidele sunt degradate de un set de enzime lizozomale care catalizează eliminarea pas cu pas a unităților de zahăr, producând în cele din urmă o ceramidă..
O altă cale de degradare constă în internalizarea sfingolipidelor în veziculele endocitice care sunt trimise înapoi la membrana plasmatică sau transportate la lizozomi unde sunt degradate de hidrolaze acide specifice..
Nu toate bazele cu lanț lung sunt reciclate, reticulul endoplasmatic are o cale pentru degradarea lor terminală. Acest mecanism de degradare constă în fosforilare în loc de acilare a LCB-urilor, dând naștere la molecule de semnalizare care pot fi substraturi solubile pentru enzimele liasa care taie LCB-fosfatul pentru a genera aciloaldehide și fosfoetanolamină..
Metabolismul acestor lipide este reglat la diferite niveluri, unul dintre ele este cel al enzimelor responsabile de sinteză, modificările lor post-translaționale și mecanismele lor alosterice..
Unele mecanisme de reglare sunt specifice celulei, fie pentru a controla momentul dezvoltării celulei în care sunt produse, fie ca răspuns la semnale specifice..



Nimeni nu a comentat acest articol încă.