
moștenirea postmendeliană Include toate ideile și / sau lucrările care au apărut după studiile publicate de naturalistul religios Gregorio Mendel în 1865 și care explică fenomene genetice sau comportamente diferite de cele descrise de acest.
Gregorio Mendel este considerat tatăl geneticii, deoarece, prin studierea încrucișărilor între plantele de mazăre, a stabilit primele legi ale moștenirii cu mai bine de 100 de ani în urmă. Prima lege, numită „Legea segregării”, propune că genele pot exista sub forme diferite sau alternative.

Aceste forme alternative se numesc „alele” și influențează direct caracteristicile fenotipice ale unei specii. Alelele pot determina culoarea părului, culoarea ochilor, tipul de păr etc..
În prima sa lege, Mendel a propus că fiecare trăsătură fenotipică transmisă de la părinți la copiii lor este controlată de două alele, fiecare provenind de la un părinte (una de la „mamă” și una de la „tată”).
Potrivit acestui lucru, atunci când părinții contribuie cu alele diferite, descendența rezultă cu un amestec, unde una dintre alele este dominantă (cea care este exprimată în fenotip) și cealaltă este recesivă (nu este văzută sau este „latentă”) ")..
A doua lege a lui Mendel este cunoscută sub numele de „Legea moștenirii independente”, întrucât în crucile sale, Mendel a observat că tiparul moștenirii care a caracterizat alelele care au definit o trăsătură nu a influențat tiparul moștenirii alelelor care a definit o trăsătură diferită.
Indice articol
De când Gregor Mendel și-a propus faimoasele sale legi cu mai bine de 100 de ani în urmă, au fost elucidate noi mecanisme pentru a explica moștenirea personajelor, dintre care multe s-ar putea spune că sunt în contradicție sau diferă considerabil de cele descrise de Mendel..
Un exemplu al fenomenelor descrise de știința „modernă”, ale căror progrese au fost posibile doar grație avansării strategiilor experimentale și a metodelor de cercetare, sunt moștenirea legată de sex, dominarea incompletă și codominanța, printre altele..
Moștenirea legată de sex are legătură cu genele care se găsesc doar pe cromozomii sexuali. La om, sexul este determinat de cromozomii X și Y. Femeile au două copii ale lui X (XX), iar bărbații au unul dintre X și unul din Y (XY)..
Genele găsite pe fiecare dintre acești doi cromozomi sunt cele care contribuie la acele trăsături care sunt legate de sex. Genele cromozomului X sunt prezente la bărbați și femei, dar genele Y sunt doar la bărbați.
Moștenirea legată de sex a fost observată și descrisă pentru prima dată de William Bateson, unul dintre primii oameni de știință care a „redescoperit” munca lui Mendel și care a profitat de acestea pentru a efectua noi experimente pe baza concluziilor sale..
Bateson a realizat cruci experimentale de găini cu penaje de diferite culori, unele cu pene întunecate și netede și altele cu pene netede și cu un model dungat..
Prin aceste încrucișări a observat că, la descendenți, masculii rezultați aveau întotdeauna un penaj cu un model dungat.
Abia în a treia generație, când un mascul dungat a fost încrucișat cu o găină cu pene întunecate și netede, s-au obținut niște cocoși cu penaj negru. Această proporție rezultată a încrucișărilor a sfidat proporțiile propuse de Mendel.
După lucrarea lui Bateson, Thomas Henry Morgan a găsit o explicație logică pentru rezultatele ciudate obținute de Bateson, atribuind diferența de colorare la găini faptului că modelul dungat a fost codificat de o alelă dominantă găsită pe cromozomul X..
La găini, spre deosebire de oameni, sexul heterogametic (XY) corespunde femelelor, în timp ce sexul homogametic corespunde bărbaților (XX).
Alela studiată de Bateson și Morgan, atunci, a fost exprimată de toți acei masculi cu penaj dungat, dar atunci când un mascul a moștenit doi cromozomi X cu alela corespunzătoare colorației negre, au apărut apoi bărbații cu penaj întunecat și fără model dungat..
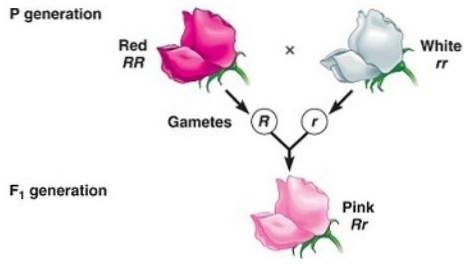
Dominația incompletă este atunci când fenotipul observat la descendenți care rezultă din încrucișarea a doi indivizi homozigoti (unul dominant și celălalt recesiv) este un fenotip „intermediar” între dominant și recesiv..
Are legătură cu „incapacitatea” alelei „dominante” de a masca complet expresia alelei recesive, motiv pentru care fenotipul rezultat corespunde unui amestec de trăsături codificate de ambele alele, de multe ori descris ca „diluat” fenotip ”De dominant.
De exemplu, în plantele de la Dondiego noaptea (Mirabilis jalapa) culoarea tipică a petalelor de flori sălbatice este roșie. Când o plantă cu petale roșii este încrucișată cu o plantă cu linie pură cu petale albe, se obține că toți descendenții sunt petale roz (roz).
Când doi indivizi homozigoti dominanți sunt încrucișați și se observă un al treilea fenotip la descendenți care corespunde celor două caracteristici determinate de cele două alele parentale, vorbim de codominanță.
În acest caz, continuând cu un exemplu similar cu cel al florilor din exemplul anterior, „al treilea fenotip” nu ar corespunde florilor roz, ci unei plante cu flori albe și flori roșii pe aceeași tulpină, din moment ce cele două alele „codomina”, Și nici unul nu-l maschează pe celălalt.
Cu toate acestea, de multe ori, codominanța și dominanța incompletă sunt dificil de distins fenotipic, deoarece ambele produc un fenotip al unui individ care pare să exprime două alele diferite..
Cu toate acestea, din punct de vedere genotipic, indivizii care prezintă alele codominante sunt diferite de cei care prezintă dominanță incompletă, întrucât într-un caz cele două alele dominante sunt exprimate în aceeași cantitate, iar în celălalt proporțiile sunt inegale..
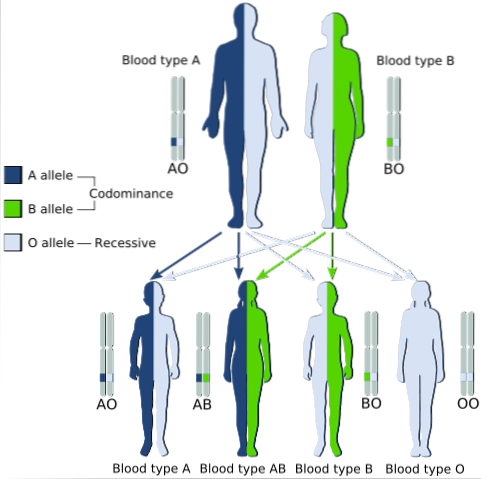
Un exemplu clasic de codominanță este tipul grupului sanguin la om, deoarece alelele care codifică antigenele A și B la populațiile umane sunt codominante.
Adică, atunci când alelele A și B se găsesc împreună în celulele unui individ, el are grupa sanguină AB.
Ele sunt alele recesive pentru un caracter care, atunci când sunt găsite în formă homozigotă, sunt letale pentru individ. Persoanele care prezintă aceste alele nu se nasc, deoarece împiedică funcționarea normală a mașinilor moleculare și împiedică translația genelor.
Multe locuri din genomul unui individ interacționează cu alte părți ale aceluiași genom, adică modificarea specifică a unei alele din genom poate condiționa direct expresia alelelor din celulele unui individ.
Aceste interacțiuni sunt dificil de determinat și sunt clasificate în două tipuri diferite: gene care interacționează în aceeași cale genetică și cele care interacționează în căi genetice diferite..
Un exemplu clasic și bine cunoscut de gene care interacționează în diferite căi genetice are legătură cu genele care determină culoarea pielii șerpilor de porumb (Pantherophis guttatus), a cărui culoare comună este un model repetitiv de pete negre și portocalii.

Acest fenotip este produs de doi pigmenți diferiți care se află sub control genetic strict. O genă determină culoarea portocalie și are două alele: una O +, care codifică pigmentul și alta O-, care determină absența totală a pigmentului portocaliu..
O altă genă diferită determină prezența sau absența pigmentului negru, alelele acestei gene sunt cunoscute sub numele de B +, cea care determină prezența pigmentului negru și B-, care determină absența pigmentului.
Ambele gene nu sunt legate și determină în mod independent culoarea șerpilor, adică un șarpe cu genotipul O + / B + este un șarpe normal cu fenotip portocaliu și negru, dar orice variație genotipică va avea ca rezultat un fenotip diferit..
Exemplul clasic al acestor gene este cel care a fost descris pentru florile „campanule” (din familia Campanulaceae), al căror fenotip „sălbatic” corespunde plantelor cu flori albastre..
Culoarea albastră a florilor albastru este determinată nu de o genă, ci de o cale metabolică în care mai multe enzime, fiecare codificată de o genă diferită, participă la sinteza unui pigment..
Astfel, există fenotipuri caracterizate prin faptul că sunt rezultatul interacțiunii mai multor gene, ale căror produse participă la aceeași cale metabolică și unde mutația sau funcționarea defectuoasă a oricăreia dintre acestea produce un fenotip, în acest caz, al florilor albe, depigmentate..



Nimeni nu a comentat acest articol încă.