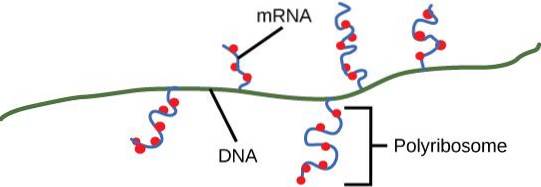
A polisom este un grup de ribozomi recrutați pentru traducerea aceluiași ARN mesager (ARNm). Structura este mai bine cunoscută sub numele de polirribozom sau ergozomul mai puțin comun..
Polizomii permit producția crescută de proteine din acei mesageri care sunt supuși traducerii simultane de mai mulți ribozomi. Polizomii participă, de asemenea, la procesele de pliere co-translațională și la achiziționarea de structuri cuaternare de către proteinele nou sintetizate..
Polizomii, împreună cu așa-numitele corpuri P și granule de stres, controlează soarta și funcția mesagerilor din celulele eucariote..
Polizomii au fost observați atât în celulele procariote, cât și în celulele eucariote. Acest lucru înseamnă că acest tip de formațiune macromoleculară are o istorie lungă în lumea celulară. Un polizom poate fi alcătuit din cel puțin doi ribozomi pe același mesager, dar în general există mai mult de doi.
În cel puțin o celulă de mamifer, pot exista până la 10.000.000 de ribozomi. S-a observat că mulți sunt liberi, dar o mare parte este asociată în polisomii cunoscuți.
Indice articol
Ribozomii tuturor ființelor vii constau din două subunități: subunitatea mică și subunitatea mare. Subunitatea mică a ribozomilor este responsabilă pentru citirea ARN-ului mesager.
Subunitatea mare este responsabilă pentru adăugarea liniară a aminoacizilor la peptida născută. O unitate translațională activă este una în care un ARNm a fost capabil să recruteze și să permită asamblarea ribozomului. După aceea, citirea tripletelor în messenger și interacțiunea cu tRNA încărcat corespunzător continuă secvențial..
Ribozomii sunt elementele de bază ale polizomilor. De fapt, ambele moduri de traducere a unui mesager pot coexista în aceeași celulă. Dacă toate componentele care alcătuiesc mașina de translație a celulei sunt purificate, am găsi patru fracții principale:
În celulele eucariote, mARN-urile sunt exportate din nucleu ca ribonucleoproteine mesager. Adică, mesagerul este cuplat cu mai multe proteine care vor determina exportul, mobilizarea și traducerea acestuia..
Printre acestea, există mai multe care interacționează cu proteina PABP atașată la coada poliA 3 'a mesagerului. Altele, cum ar fi complexul CBP20 / CBP80, se vor lega de capota 5 'a ARNm.
Eliberarea complexului CBP20 / CBP80 și recrutarea subunităților ribozomale pe capota 5 'definesc formarea ribozomului.
Traducerea începe și noi ribozomi sunt asamblate pe capota 5 '. Acest lucru se întâmplă pentru un număr limitat de ori care depinde de fiecare mesager și de tipul de polisom în cauză..
După această etapă, factorii de alungire a traducerii asociați cu capacul la capătul 5 'interacționează cu proteina PABP legată de capătul 3' al ARNm. Se formează astfel un cerc definit prin unirea regiunilor netraductibile ale mesagerului. Astfel, sunt recrutați cât mai mulți ribozomi pe măsură ce permite lungimea mesagerului și alți factori.

Alți polizomi pot adopta o configurație liniară dublă sau spirală cu patru ribozomi pe tură. Forma circulară a fost asociată cel mai puternic cu polizomii liberi.
Polizomii se formează pe unități de translație active (inițial monozomi) cu adăugarea secvențială a altor ribozomi pe același ARNm.
În funcție de localizarea lor subcelulară, găsim trei tipuri diferite de polizomi, fiecare având funcțiile sale specifice..
Se găsesc liberi în citoplasmă, fără asocieri aparente cu alte structuri. Acești polizomi traduc ARNm care codifică proteinele citosolice.
Deoarece învelișul nuclear este o extensie a reticulului endoplasmatic, acest tip de polizom poate fi asociat și cu învelișul nuclear exterior.
În acești polizomi, ARNm-urile care codifică două grupuri importante de proteine sunt traduse. Unele, care sunt o parte structurală a reticulului endoplasmatic sau a complexului Golgi. Altele, care trebuie modificate post-translațional și / sau mutate intracelular de către aceste organite.
Polizomii asociați cito-scheletului traduc proteinele din ARNm care sunt concentrate asimetric în anumite compartimente subcelulare.
Adică, la ieșirea din nucleu, unele ribonucleoproteine mesager sunt mobilizate către locul unde este necesar produsul pe care îl codifică. Această mobilizare este realizată de citoschelet cu participarea proteinelor care se leagă de coada poliA a ARNm..
Cu alte cuvinte, citoscheletul distribuie mesagerii după destinație. Această soartă este indicată de funcția proteinei și de locul în care trebuie să locuiască sau să acționeze..
Chiar dacă un ARNm este transcris, asta nu înseamnă neapărat că trebuie tradus. Dacă acest ARNm este degradat în mod specific în citoplasma celulară, se spune că expresia genei sale este reglată post-transcripțional..
Există multe modalități de a realiza acest lucru, iar una dintre ele este prin acțiunea așa-numitelor gene MIR. Produsul final al transcrierii unei gene MIR este un microARN (miARN).
Acestea sunt complementare sau parțial complementare cu alți mesageri a căror traducere o reglează (mutarea post-transcripțională). Tăcerea poate implica și degradarea specifică a unui anumit mesager.
Tot ceea ce ține de traducere, compartimentarea, reglarea și mutarea genei post-transcripționale este controlat de polizomi..
Pentru a face acest lucru, ei interacționează cu alte macrostructuri moleculare ale celulei cunoscute sub numele de corpuri P și granule de stres. Acești trei corpuri, ARNm și microARN, definesc astfel proteomul prezent într-o celulă la un moment dat..


Nimeni nu a comentat acest articol încă.