
dihibridism, În genetică, definește studiul simultan a două caractere ereditare diferite și, prin extensie, a celor a căror manifestare depinde de două gene diferite, chiar dacă este același caracter.
Cele șapte trăsături pe care le-a analizat Mendel i-au fost utile în prezentarea teoriei sale despre moștenirea caracterelor deoarece, printre altele, genele responsabile de manifestarea lor aveau alele contrastante al căror fenotip era ușor de analizat și pentru că fiecare a determinat expresia unică caracter.
Adică au fost trăsături monogene a căror stare hibridă (monohibrizi) a permis determinarea relațiilor de dominanță / recesivitate între alelele acelei gene..
Când Mendel a analizat moștenirea comună a două personaje diferite, a procedat așa cum a făcut-o cu personajele singure. El a obținut hibrizi dubli (dihidri) care i-au permis să verifice:
Știm acum că moștenirea personajelor este puțin mai complexă decât ceea ce a observat Mendel, dar și că, în fundamentele sale, Mendel a fost complet corect..
Dezvoltarea ulterioară a geneticii a făcut posibilă demonstrarea faptului că încrucișările dihidri și analiza lor (dihidridismul), așa cum a putut Bateson să demonstreze inițial, ar putea fi o sursă inepuizabilă de descoperiri în această știință puternică și naștentă a secolului XX..
Datorită utilizării lor inteligente, au reușit să dea geneticianului o idee oarecum mai clară despre comportamentul și natura genelor..
Indice articol
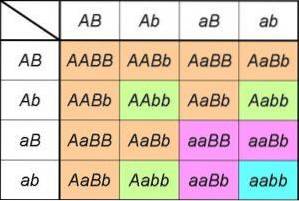
Dacă analizăm produsele unei cruci monohibridă Aa X Aa, putem observa că este egal cu dezvoltarea produsului remarcabil (LA+la)Două= AA + 2Aa + aa.
Expresia din stânga include cele două tipuri de gamete pe care le poate produce unul dintre părinții heterozigoți pentru genă LA/la; prin pătrat indicăm că ambii părinți sunt de constituție identică pentru gena studiată [adică este o cruce monohibridă (LA+la) X (LA+la)]].
Expresia din dreapta ne oferă genotipurile (și, prin urmare, fenotipurile sunt deduse) și proporțiile așteptate derivate din încrucișare.
Prin urmare, putem observa direct proporțiile genotipice derivate din prima lege (1: 2: 1), precum și proporțiile fenotipice explicate de aceasta (1AA+DouăAa= 3LA_ pentru fiecare 1aa, sau raport fenotipic 3: 1).
Dacă luăm acum în considerare o cruce pentru a analiza moștenirea unei gene B, expresiile și proporțiile vor fi aceleași; de fapt, va fi așa pentru orice genă. Prin urmare, într-o cruce dihidridă, avem de fapt dezvoltarea produselor din (LA+la)Două X (B+b)Două.
Sau ce este la fel, dacă crucea dihidridă implică două gene care participă la moștenirea a două caractere fără legătură, proporțiile fenotipice vor fi cele prezise de a doua lege: (3LA_: 1aa) X (3B_: 1bebelus) = 9LA_B_: 3LA_bebelus: 3aaB_: 1aabb).
Acestea, desigur, sunt derivate din rapoartele de genotip ordonate 4: 2: 2: 2: 2: 1: 1: 1: 1 rezultate din produsul de (LA+la)Două X (B+b)Două= (AA + 2Aa + aa) X (BB + DouăBb + bebelus).
Vă invităm să vedeți singur pentru a analiza acum ce se întâmplă atunci când raporturile fenotipice 9: 3: 3: 1 ale unei cruci dihybrid „deviază” de la aceste relații matematice clare și previzibile care explică moștenirea independentă a două caractere codificate. diferite gene.
Există două modalități principale prin care încrucișările dihidri se abată de la ceea ce este „așteptat”. Primul este cel în care analizăm moștenirea comună a două caractere diferite, dar proporțiile fenotipice observate în descendenți dau o predominanță clară manifestării fenotipurilor parentale..
Cel mai probabil este un caz de gene legate. Adică, cele două gene analizate, deși sunt la locuri diferite, sunt atât de apropiate unele de altele fizic încât tind să fie moștenite împreună și, evident, nu sunt distribuite independent..
Cealaltă circumstanță, care este, de asemenea, destul de comună, derivă din faptul că o mică minoritate a trăsăturilor ereditare sunt monogene..
Dimpotrivă, mai mult de două gene participă la manifestarea celor mai multe trăsături ereditare..
Din acest motiv, este întotdeauna posibil ca interacțiunile genetice stabilite între genele care participă la manifestarea unui singur caracter să fie complexe și să depășească o simplă relație de dominanță sau recesivitate, așa cum se observă în relațiile alelice tipice trăsăturilor monogene..
De exemplu, în manifestarea unei trăsături, aproximativ patru enzime pot participa într-o anumită ordine pentru a da naștere produsului final responsabil pentru manifestarea fenotipică a fenotipului sălbatic..
Analiza care face posibilă identificarea numărului de gene din diferiți loci care participă la manifestarea unei trăsături genetice, precum și ordinea în care acționează, se numește analiză epistazică și este probabil cea care definește cel mai de obicei ceea ce noi numiți analiză genetică.în sensul său cel mai clasic.
La sfârșitul acestui post, sunt prezentate proporțiile fenotipice observate în cele mai frecvente cazuri de epistază - și acest lucru ținând cont doar de încrucișările dihidri..
Prin creșterea numărului de gene care participă la manifestarea aceleiași trăsături, complexitatea interacțiunilor genetice și interpretarea lor crește evident..
În plus, care la rândul său poate fi luată ca regulă de aur pentru un diagnostic corect al interacțiunilor epistatice, poate fi verificată apariția de noi fenotipuri care nu sunt prezente în generația părintească..
În cele din urmă, în afară de a permite analiza apariției unor noi fenotipuri și a proporției acestora, analiza epistazei permite, de asemenea, determinarea ordinii ierarhice în care diferitele gene și produsele lor trebuie să se manifeste într-o cale determinată pentru a explica fenotipul asociat acestora..
Gena de manifestare cea mai de bază sau timpurie este epistatică peste toate celelalte, deoarece fără produsul sau acțiunea ei, de exemplu, cei din aval de ea nu vor putea să se exprime, ceea ce va fi, prin urmare, ipostatic pentru ea..
O genă / produs aflat pe locul trei în ierarhie va fi ipostatică pentru primele două și epistatică pentru oricare altele care rămân în această cale de expresie genică..



Nimeni nu a comentat acest articol încă.