
aparate Golgi, De asemenea, cunoscut sub numele de complex Golgi, este un organit de celule membranare format dintr-un set de vezicule plate stivuite împreună; Aceste pungi au fluid în interior. Se găsește într-o mare varietate de eucariote, inclusiv animale, plante și ciuperci.
Acest organet este responsabil pentru prelucrarea, ambalarea, clasificarea, distribuția și modificarea proteinelor. În plus, are și un rol în sinteza lipidelor și carbohidraților. Pe de altă parte, în plante, sinteza componentelor peretelui celular are loc în aparatul Golgi..
Aparatul Golgi a fost descoperit în 1888, în timp ce celulele nervoase erau studiate; descoperitorul său, Camillo Golgi, a câștigat Premiul Nobel. Structura este detectabilă prin colorarea cromatului de argint.
La început existența organului a fost îndoielnică pentru oamenii de știință de atunci și au atribuit observațiile Golgi unor artefacte simple produse ale tehnicilor utilizate.
Indice articol
Aparatul Golgi este un organet eucariot de natură membrană. Seamănă cu pungile în stive, deși organizarea poate varia în funcție de tipul celulei și de organism. Este responsabil pentru modificarea proteinelor după traducere.
De exemplu, unii carbohidrați pot fi adăugați pentru a forma o glicoproteină. Acest produs este ambalat și distribuit în compartimentul celular unde este necesar, cum ar fi membrana, lizozomii sau vacuolele; poate fi trimis și în afara celulei. De asemenea, participă la sinteza biomoleculelor.
Citoscheletul (în mod specific actina) determină localizarea acestuia și, în general, complexul este situat într-o zonă a interiorului celulei aproape de nucleu și centrosom.
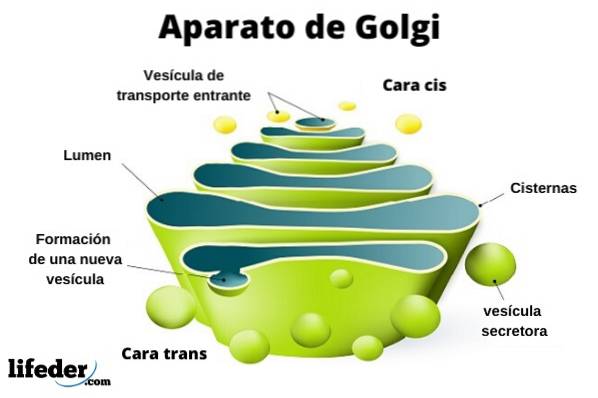
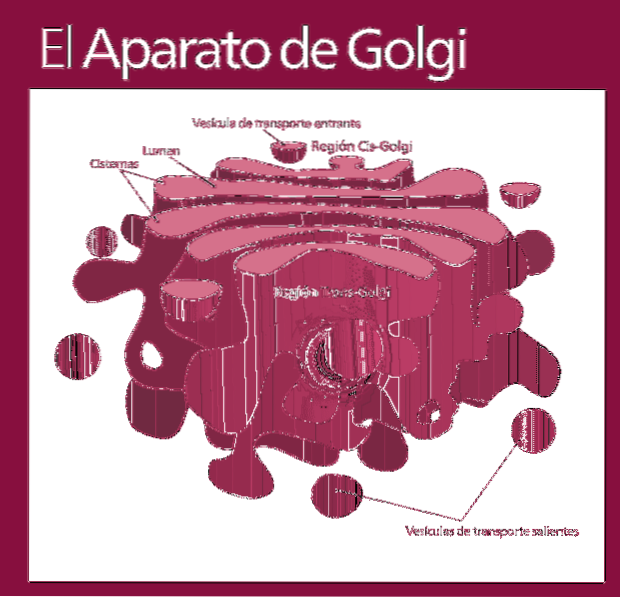
Complexul Golgi este alcătuit dintr-un set de pungi plate, fenestrate, în formă de disc, numite cisterne Golgiene, cu grosime variabilă.
Aceste saci sunt stivuite, în grupuri de patru sau șase cisterne. Într-o celulă de mamifer puteți găsi între 40 și 100 de baterii conectate între ele.
Complexul Golgi prezintă o caracteristică interesantă: există polaritate în ceea ce privește structura și, de asemenea, funcțional.
Puteți distinge între fața cis și fața trans. Primul este legat de intrarea proteinelor și este situat în apropierea reticulului endoplasmatic. A doua este fața de ieșire sau secreția produsului; Sunt formate din unul sau două tancuri care au o formă tubulară.
Alături de această structură sunt vezicule care alcătuiesc sistemul de transport. Mormanele de saci sunt atașate între ele într-o structură care amintește de un arc sau curmale.
La mamifere, complexul Golgi este fragmentat în mai multe vezicule în timpul proceselor de diviziune celulară. Veziculele trec la celulele fiice și iau din nou forma tradițională a complexului.
Organizarea complexului nu este comună în toate grupurile de organisme. În unele tipuri de celule complexul nu este structurat ca seturi de cisterne stivuite în grupuri; dimpotrivă, sunt localizate individual. Un exemplu al acestei organizații este cel al ciupercii Saccharomyces cerevisiae.
La unele organisme unicelulare, cum ar fi toxoplasma sau tripanosom, a fost raportată prezența unui singur teanc membranos.
Toate aceste excepții indică faptul că stivuirea structurilor nu este esențială pentru îndeplinirea funcției lor, deși apropierea dintre saci face procesul de transport mult mai eficient..
În mod similar, unor eucariote bazale le lipsește aceste cisterne; de exemplu, ciupercile. Aceste dovezi susțin teoria conform căreia aparatul a apărut într-o descendență mai târzie decât primele eucariote..
Funcțional, complexul Golgi este împărțit în următoarele compartimente: rețeaua cis, pungile stivuite - care la rândul lor sunt împărțite în subcompartimentul mediu și trans - și trans.
Moleculele care vor fi modificate intră în complexul Golgi urmând aceeași ordine (rețea cis, urmată de subcompartimente care vor fi excretate în final în rețeaua trans).
Cele mai multe reacții apar în zonele cele mai active: subcompartimentele trans și mijlocii.
Complexul Golgi are ca funcție principală modificarea post-translațională a proteinelor datorită enzimelor pe care le au în interior.
Aceste modificări includ procese de glicozilare (adăugarea de carbohidrați), fosforilare (adăugarea unei grupări fosfat), sulfatare (adăugarea unei grupări fosfat) și proteoliză (degradarea proteinelor).
Mai mult, complexul Golgi este implicat în sinteza biomoleculelor specifice. Fiecare dintre funcțiile sale este descrisă în detaliu mai jos:
În aparatul Golgi apare modificarea unei proteine la o glicoproteină. PH-ul acid tipic din interiorul organitei este esențial pentru ca acest proces să aibă loc în mod normal..
Există un schimb constant de materiale între aparatul Golgi cu reticulul endoplasmatic și lizozomii. În reticulul endoplasmatic, proteinele suferă și modificări; acestea includ adăugarea unei oligozaharide.
Când aceste molecule (N-oligozaharide) intră în complexul Golgi, primesc o serie de modificări suplimentare. Dacă destinul acestei molecule urmează să fie transportat în afara celulei sau să fie primit în membrana plasmatică, apar modificări speciale.
Aceste modificări includ următoarele etape: îndepărtarea a trei reziduuri de manoză, adăugarea de N-acetilglucozamină, îndepărtarea a două manoze și adăugarea de fucoză, două N-acetilglucozamină suplimentare, trei galactoză și trei reziduuri de acid sialic..
În schimb, proteinele care sunt destinate lizozomilor sunt modificate în felul următor: nu există eliminarea manozelor ca etapă inițială; în schimb, are loc fosforilarea acestor reziduuri. Acest pas are loc în regiunea cis a complexului.
Apoi, grupurile N-acetilglucozamină sunt îndepărtate lăsând manozele cu fosfat adăugat la oligozaharidă. Acești fosfați indică faptul că proteina trebuie să fie direcționată în mod specific către lizozomi..
Receptorii care sunt responsabili de recunoașterea fosfaților care indică soarta lor intracelulară se află în rețeaua trans.
În complexul Golgi apare sinteza glicolipidelor și sfingomielinului, folosind ceramida (sintetizată anterior în reticulul endoplasmatic) ca moleculă de origine. Acest proces este contrar celui al restului de fosfolipide care alcătuiesc membrana plasmatică, care sunt derivate din glicerol..
Sfingomielina este o clasă de sfingolipide. Este o componentă abundentă a membranelor mamiferelor, în special a celulelor nervoase, unde fac parte din teaca de mielină.
După sinteza lor, sunt transportați la locația lor finală: membrana plasmatică. Capetele lor polare sunt situate spre exteriorul suprafeței celulei; aceste elemente au un rol specific în procesele de recunoaștere a celulelor.
În celulele vegetale, aparatul Golgi contribuie la sinteza polizaharidelor care alcătuiesc peretele celular, în special hemiceluloză și pectine. Prin transport vezicular, acești polimeri sunt transportați în afara celulei.
La legume, acest pas este crucial și aproximativ 80% din activitatea reticulului este alocată sintezei polizaharidelor. De fapt, sute de organite au fost raportate în celulele vegetale..
Diferitele biomolecule - proteine, carbohidrați și lipide - sunt transferate către destinațiile lor celulare de către complexul Golgi. Proteinele au un fel de „cod” care este responsabil pentru informarea destinației căreia îi aparține.
Acestea sunt transportate în vezicule care părăsesc rețeaua trans și se deplasează către compartimentul celular specific.
Proteinele pot fi transportate la membrană printr-o cale constitutivă specifică. De aceea există o încorporare continuă de proteine și lipide în membrana plasmatică. Proteinele a căror destinație finală este complexul Golgi sunt reținute de aceasta.
În plus față de calea constitutivă, alte proteine sunt destinate exteriorului celulei și apar prin semnale din mediu, fie că sunt hormoni, enzime sau neurotransmițători..
De exemplu, în celulele pancreatice, enzimele digestive sunt ambalate în vezicule care sunt secretate numai atunci când este detectată prezența alimentelor..
Cercetări recente raportează existența unor căi alternative pentru proteinele de membrană care nu trec prin aparatul Golgi. Cu toate acestea, aceste căi ocolire „Neconvenționale” sunt dezbătute în literatură.
Există cinci modele pentru a explica traficul de proteine în aparat. Primul implică traficul de material între compartimente stabile, fiecare având enzimele necesare pentru a îndeplini funcții specifice. Al doilea model implică maturarea cisternelor progresiv.
Al treilea propune și maturarea pungilor, dar cu încorporarea unei noi componente: transportul tubular. Conform modelului, tubulii sunt importanți în trafic în ambele sensuri.
Al patrulea model propune ca complexul să funcționeze ca o unitate. Al cincilea și ultimul model este cel mai recent și susține că complexul este împărțit în diferite compartimente.
În anumite tipuri de celule complexul Golgi are funcții specifice. Celulele pancreasului au structuri specializate pentru secreția de insulină.
Diferitele tipuri de sânge la om sunt un exemplu de modele de glicozilare diferențială. Acest fenomen se explică prin prezența diferitelor alele care codifică glucotransferaza.


Nimeni nu a comentat acest articol încă.