eritropoietină, hemopoietină sau EPO este o glicoproteină cu funcții hormonale (citokine) responsabilă pentru controlul proliferării, diferențierii și supraviețuirii celulelor progenitoare ale eritrocitelor sau celulelor roșii din sânge în măduva osoasă, adică a eritropoiezei.
Această proteină este unul dintre diferiții factori de creștere care controlează procesele hematopoietice prin care, dintr-un grup mic de celule stem pluripotente, se formează celulele găsite în sânge: atât eritrocite, cât și globule albe și limfocite. Adică, celulele descendenței mieloide și limfoide.
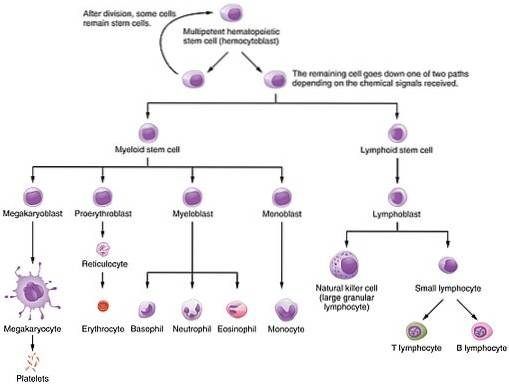
Importanța sa constă în importanța funcțională a celulelor care ajută la înmulțire, diferențiere și maturare, deoarece eritrocitele sunt responsabile pentru transportul oxigenului din plămâni către diferitele țesuturi ale corpului.
Eritropoietina a fost primul factor de creștere care a fost clonat (în 1985), iar administrarea sa pentru tratamentul cu succes al anemiei cauzate de insuficiența renală este aprobată în prezent de Administrația Americană pentru Alimente și Medicamente (FDA)..
Noțiunea că eritropoieza este controlată de un factor umoral (factor solubil prezent în circulație) a fost propusă în urmă cu mai bine de 100 de ani de către Carnot și Deflandre atunci când au studiat efectele pozitive asupra creșterii procentelor de celule roșii la iepurii tratați cu ser. animale.
Cu toate acestea, abia în 1948 Bonsdorff și Jalavisto au introdus termenul „eritropoietină” pentru a descrie factorul umoral cu o implicație specifică asupra producției de eritrocite..
Indice articol
Eritropoietina este o proteină din familia glicoproteinelor. Este stabil la pH acid și are o greutate moleculară de aproximativ 34 kDa..
Are aproximativ 193 de aminoacizi, inclusiv o regiune N-terminală hidrofobă cu 27 de reziduuri, care este îndepărtată prin prelucrare co-translațională; și un reziduu de arginină la poziția 166 care se pierde și el, deci proteina circulantă are 165 de aminoacizi.
În structura sa, se poate observa formarea a două punți disulfură între reziduurile de cisteină prezente în pozițiile 7-161 și 29-33, care sunt legate de funcționarea sa. Este compus mai mult sau mai puțin de 50% din helice alfa, care aparent participă la formarea unei regiuni sau porțiuni globulare.
Are 40% carbohidrați, reprezentat de trei lanțuri oligozaharidice legate de N la diferite reziduuri de acid aspartic (Asp) și un lanț O legat de un reziduu de serină (Ser). Aceste oligozaharide sunt compuse în principal din fucoză, manoză, N-acetil glucozamină, galactoză și acid N-acetil neuraminic..
Regiunea glucidică a EPO îndeplinește mai multe roluri:
- Este esențial pentru activitatea sa biologică.
- O protejează de degradarea sau deteriorarea cauzată de radicalii liberi de oxigen.
- Lanțurile oligozaharidice sunt necesare pentru secreția proteinei mature.
La om, gena care codifică această proteină se află în mijlocul brațului lung al cromozomului 7, în regiunea q11-q22; se găsește într-o singură copie într-o regiune de 5,4 kb și are cinci exoni și patru introni. Studiile de omologie indică faptul că secvența sa are o identitate de 92% cu cea a altor primate și 80% cu cea a unor rozătoare..
În timpul dezvoltării fetale, eritropoietina este produsă în principal în ficat, dar s-a stabilit că, în aceeași etapă, gena care codifică acest hormon este, de asemenea, exprimată abundent în regiunea mijlocie a nefronilor renali..
După naștere, în ceea ce ar putea fi considerat toate etapele postnatale, hormonul este produs în esență în rinichi. Mai exact, de către celulele cortexului și suprafața corpusculilor renali.
Ficatul participă, de asemenea, la producerea de eritropoietină în etapele postnatale, din care se excretă aproximativ 20% din conținutul total de EPO circulant..
Alte organe „extra-renale” în care a fost detectată producția de eritropoietină includ celule endoteliale periferice, celule musculare netede vasculare și celule producătoare de insulină..
Unele centre de secreție EPO sunt, de asemenea, cunoscute că există în sistemul nervos central, inclusiv hipocampul, cortexul, celulele endoteliale ale creierului și astrocitele..
Producția de eritropoietină nu este controlată direct de numărul de globule roșii din sânge, ci de aportul de oxigen din țesuturi. O deficiență de oxigen în țesuturi stimulează producția de EPO și receptorii săi în ficat și rinichi.
Această activare a expresiei genice mediată de hipoxie este produsul activării căii unei familii de factori de transcripție cunoscuți ca factor 1 inductibil de hipoxie (HIF-1). factor 1 inductibil de hipoxie).
Hipoxia, atunci, induce formarea multor complexe proteice care îndeplinesc diferite funcții în activarea expresiei eritropoietinei și care se leagă direct sau indirect de factori care traduc semnalul de activare către promotorul genei EPO, stimulând transcrierea acesteia..
Alți factori de stres, cum ar fi hipoglicemia (glicemia scăzută), creșterea calciului intracelular sau prezența speciilor reactive de oxigen, declanșează, de asemenea, calea HIF-1..
Mecanismul de acțiune al eritropoietinei este destul de complex și depinde în principal de capacitatea sa de a stimula diferite cascade de semnalizare implicate în proliferarea celulară, care sunt legate, la rândul lor, de activarea altor factori și hormoni..
În corpul uman al unui adult sănătos există un echilibru între producerea și distrugerea globulelor roșii sau a eritrocitelor, iar EPO participă la menținerea acestui echilibru prin înlocuirea eritrocitelor care dispar.
Atunci când cantitatea de oxigen disponibilă în țesuturi este foarte mică, expresia genei care codifică eritropoietina crește în rinichi și ficat. Stimulul poate fi dat și de altitudini mari, hemoliză, condiții de anemie severă, sângerări sau expunere prelungită la monoxid de carbon.
Aceste condiții generează o stare de hipoxie, ceea ce determină creșterea secreției de EPO, se produc un număr mai mare de celule roșii și crește și fracțiunea de reticulocite în circulație, care sunt una dintre celulele progenitoare ale eritrocitelor..
În eritropoieză, EPO este implicată în principal în proliferarea și diferențierea celulelor progenitoare implicate în descendența globulelor roșii (progenitori eritrocitari), dar activează și mitoza în proeritroblasti și eritroblasti bazofili și, de asemenea, accelerează eliberarea reticulocitelor măduvei osoase.
Primul nivel la care funcționează proteina este în prevenirea morții celulare programate (apoptoză) a celulelor precursoare formate în măduva osoasă, pe care o atinge prin interacțiune inhibitoare cu factorii implicați în acest proces..
Celulele care răspund la eritropoietină posedă un receptor specific pentru eritropoietină cunoscut sub numele de receptor de eritropoietină sau EpoR. Odată ce proteina formează un complex cu receptorul său, semnalul este transferat în interiorul celulei: către nucleu.
Primul pas pentru transferul semnalului este o schimbare conformațională care are loc după ce proteina se leagă de receptorul său, care este, în același timp, legat de alte molecule de receptor care sunt activate. Printre acestea se numără Janus-tirozin kinaza 2 (Jack-2).
Printre unele dintre căile care sunt activate în aval, după ce Jack-2 mediază fosforilarea reziduurilor de tirozină ale receptorului EpoR, se află calea MAP kinază și proteina kinază C, care activează factorii de transcripție care cresc expresia genelor specifice.
La fel ca mulți factori hormonali din organisme, eritropoietina nu este limitată la o singură funcție. Acest lucru a fost elucidat prin numeroase investigații.
Pe lângă faptul că acționează ca factor de proliferare și diferențiere a eritrocitelor, care sunt esențiale pentru transportul gazelor prin fluxul sanguin, eritropoietina pare să îndeplinească unele funcții suplimentare, nu neapărat legate de activarea proliferării și diferențierii celulare.
Studiile au sugerat că EPO previne deteriorarea celulelor și, deși mecanismele sale de acțiune nu sunt cunoscute exact, se crede că poate preveni procesele apoptotice produse de tensiunea redusă sau absentă a oxigenului, excită toxicitatea și expunerea la radicalii liberi..
Participarea sa la prevenirea apoptozei a fost studiată prin interacțiunea cu factori determinanți în cascadele de semnalizare: Janus-tirozin kinază 2 (Jak2), caspază 9, caspază 1 și caspază 3, glicogen sintază kinază-3β, factor de activare a proteazelor apoptotice 1 (Apaf-1) și altele.
Participă la inhibarea inflamației celulare prin inhibarea unor citokine pro-inflamatorii, cum ar fi interleukina 6 (IL-6), factorul de necroză tumorală alfa (TNF-α) și proteina chimio-atractivă monocită 1.
În sistemul vascular, s-a demonstrat că colaborează la menținerea integrității sale și la formarea de noi capilare din vasele existente în zone fără vasculatură (angiogeneză). În plus, previne permeabilitatea barierei hematoencefalice în timpul leziunilor.
Se crede că stimulează neovascularizarea postnatală prin creșterea mobilizării celulelor progenitoare de la măduva osoasă la restul corpului..
Acesta joacă un rol important în dezvoltarea celulelor progenitoare neuronale prin activarea factorului nuclear KB, care promovează producția de celule stem nervoase..
Acționând în colaborare cu alte citokine, EPO are o funcție "modulantă" în controlul căilor de proliferare și diferențiere a megacariocitelor și a granulocitelor-monocite..

Nimeni nu a comentat acest articol încă.