fosfatidiletanolamină (PE) este un glicerofosfolipid abundent în membranele plasmatice ale organismelor procariote. Dimpotrivă, în membranele celulare eucariote acesta este al doilea glicerofosfolipid cel mai abundent din interiorul membranei plasmatice după fosfatidilcolină..
În ciuda abundenței fosfatidiletanolaminei, abundența acesteia depinde nu numai de tipul celulei, ci și de compartiment și de momentul ciclului de viață celular specific..
Membranele biologice sunt bariere care definesc organismele celulare. Nu numai că au funcții de protecție și izolare, dar sunt, de asemenea, cheia stabilirii proteinelor care necesită un mediu hidrofob pentru funcționarea lor optimă..
Atât eucariotele, cât și procariotele au membrane compuse în principal din glicerofosfolipide și într-o măsură mai mică sfingolipide și steroli..
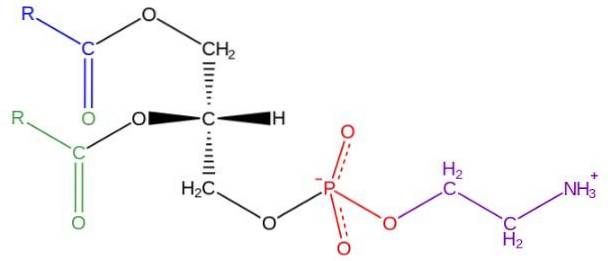
Glicerofosfolipidele sunt molecule amfipatice structurate pe o coloană vertebrală L-glicerol care este esterificată la pozițiile sn-1 și sn-2 de doi acizi grași cu lungime și grad de saturație variabile. În poziția hidroxil a poziției sn-3 este esterificat de o grupare fosfat, la care la rândul său pot fi atașate diferite tipuri de molecule care dau naștere diferitelor clase de glicerofosfolipide.
În lumea celulară există o mare varietate de glicerofosfolipide, cu toate acestea, cele mai abundente sunt fosfatidilcolină (PC), fosfatidiletanolamină (PE), fosfatidilserină (PS), fosfatidilinozitol (PI), acid fosfatidic (PA), fosfatidilglicerol (PG) și cardiol (CL).
Indice articol
Structura fosfatidiletanolaminei a fost descoperită de Baer și colab. În 1952. După cum s-a determinat experimental pentru toate glicerofosfolipidele, fosfatidiletanolamina este formată dintr-o moleculă de glicerol esterificată la pozițiile sn-1 și sn-2 cu lanțuri acide grase cu între 16 și 20 atomi de carbon.
Acizii grași esterificați în hidroxil sn-1 sunt în general saturați (fără legături duble) cu lungimi maxime de 18 atomi de carbon, în timp ce lanțurile legate în poziția sn-2 sunt mai lungi și cu una sau mai multe nesaturări (duble legături).
Gradul de saturație al acestor lanțuri contribuie la elasticitatea membranei, care are o mare influență asupra inserției și sechestrării proteinelor în stratul bistrat..
Fosfatidiletanolamina este considerată glicerofosfolipidă non-lamelară, deoarece are o formă geometrică conică. Această formă este dată de dimensiunea redusă a grupului său polar sau a „capului”, în raport cu cea a lanțurilor de acizi grași care cuprind „cozile” hidrofobe..
„Capul” sau grupul polar al fosfatidiletanolaminei are un caracter zwitterionic, adică are grupuri care pot fi încărcate pozitiv și negativ în anumite condiții de pH..
Această caracteristică îi permite legarea de hidrogen cu un număr mare de reziduuri de aminoacizi, iar distribuția sarcinii este un factor esențial pentru topologia domeniului multor proteine de membrană integrale..
În celulele eucariote sinteza lipidelor structurale este restricționată geografic, principalul situs de biosinteză fiind reticulul endoplasmatic (ER) și într-o măsură mai mică aparatul Golgi..
Există patru căi biosintetice independente pentru producerea fosfatidiletanolaminei: (1) calea CDP-etanolamină, cunoscută și sub numele de calea Kennedy; (2) calea PSD pentru decarboxilarea fosfatidilserinei (PS); (3) acilarea lizoz-PE și (4) reacții de modificare a bazei grupului polar al altor glicerofosfolipide.
Biosinteza fosfatidiletanolaminei pe această cale este limitată la ER și s-a demonstrat că în celulele hepatice de hamster este principala cale de producție. Se compune din trei etape enzimatice consecutive catalizate de trei enzime diferite.
În prima etapă, fosfoetanolamina și ADP sunt produse datorită acțiunii etanolamin kinazei, care catalizează fosforilarea ATP-dependentă a etanolaminei..
Spre deosebire de plante, nici mamiferele, nici drojdiile nu sunt capabile să producă acest substrat, deci trebuie consumat în dietă sau obținut din degradarea moleculelor de fosfatidiletanolamină sau sfingozină preexistente..
Fosfoetanolamina este utilizată de CTP: fosfoetanolamină citidiltransferază (ET) pentru a forma compusul cu energie ridicată CDP: etanolamină și un fosfat anorganic.
1,2-Diacilglicerol etanolamină fosfotransferază (ETP) folosește energia conținută în legătura CDP-etanolamină pentru a lega covalent etanolamina de o moleculă de diacilglicerol inserată prin membrană, dând naștere la fosfatidiletanolamină..
Această rută funcționează atât la procariote, cât și la drojdie și mamifere. La bacterii apare în membrana plasmatică, dar la eucariote apare într-o zonă a reticulului endoplasmatic care este strâns legată de membrana mitocondrială.
La mamifere calea este catalizată de o singură enzimă, fosfatidilserina decarboxilaza (PSD1p), care este încorporată în membrana mitocondrială, a cărei genă este codificată de nucleu. Reacția implică decarboxilarea PS la fosfatidiletanolamină.
Celelalte două căi (acilarea PE-liză și schimbul de calciu dependent de grupul polar) apar în reticulul endoplasmatic, dar nu contribuie semnificativ la producția totală de fosfatidiletanolamină în celulele eucariote..
Glicerofosfolipidele au trei funcții principale în celulă, printre care se remarcă funcțiile structurale, stocarea energiei și semnalizarea celulară..
Fosfatidiletanolamina este asociată cu ancorarea, stabilizarea și plierea mai multor proteine de membrană, precum și modificările conformaționale necesare funcționării multor enzime.
Există dovezi experimentale care propun fosfatidiletanolamina ca un glicerofosfolipid crucial în stadiul târziu al telofazei, în timpul formării inelului contractil și a stabilirii fragmoplastului care permite divizarea membranei celor două celule fiice..
De asemenea, are un rol important în toate procesele de fuziune și fisiune (unire și separare) ale membranelor atât ale reticulului endoplasmatic, cât și ale aparatului Golgi..
În E. coli s-a demonstrat că fosfatidiletanolamina este necesară pentru plierea și funcționarea corectă a enzimei lactoză permează, motiv pentru care s-a sugerat că joacă un rol ca „chaperonă” moleculară..
Fosfatidiletanolamina este principalul donator al moleculei de etanolamină necesară modificării post-translaționale a numeroaselor proteine, cum ar fi ancorele GPI.
Acest glicerofosfolipid este precursorul a numeroase molecule cu activitate enzimatică. În plus, moleculele derivate din metabolismul său, precum și diacilglicerolul, acidul fosfatidic și unii acizi grași, pot acționa ca al doilea mesager. În plus, este un substrat important pentru producerea de fosfatidilcolină..



Nimeni nu a comentat acest articol încă.